Welcome to CropSci Publisher
 Research Insights into Genetic Improvement for...
Research Insights into Genetic Improvement for...- A Review of Cotton Cultivation Techniques for ...
- Utilizing Genome Sequencing for Enhanced Pest ...
- Advances in Haploid Breeding Techniques for Ma...
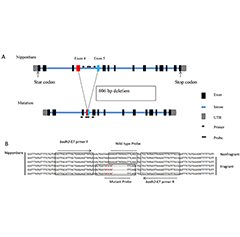
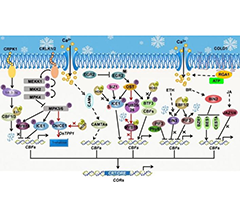
- Functional Genomics Insights into Gene Regulat...
- Meta-Analysis of Epigenetic Marks Influencing ...
- Functional Genomics of Rice: Recent Discoverie...
- Meta-analysis of Flood Tolerance Genes in Rice...
- Nodulation begins with the initiation of infect...(22 Mar.2024)
- Auxin plays a pivotal role in the co-evolution of pl...(22 Mar.2024)
- Auxin plays a pivotal role in the co-evolution of pl...(21 Mar.2024)
- Polyploidization is a major driver of genome diversi...(21 Mar.2024)