Research Article
Identification of Loss of Rf1 Gene Function Phenomenon in Restorer Line and Its Mutant of Dian-Type Hybrid Rice 
2 State Key Laboratory for Conservation and Utilization of Bio-Resources in Yunnan, Yunnan Agricultural University, Kunming, 650201, P.R. China
* These authors contributed equally to this work
 Author
Author  Correspondence author
Correspondence author
Rice Genomics and Genetics, 2021, Vol. 12, No. 4 doi: 10.5376/rgg.2021.12.0004
Received: 10 Aug., 2021 Accepted: 21 Aug., 2021 Published: 11 Sep., 2021
Feng T., Zhu Q., Guo X.Q., Li W., Yu W.L., Li S.C., Wu W.B., Wan Z.W., Huang D.J., Chen L.J., and Lee D.S., 2021, Identification of loss of Rf1 gene function phenomenon in restorer line and its mutant of Dian-type hybrid rice, Rice Genomics and Genetics, 12(4): 1-8 (doi: 10.5376/rgg.2021.12.0004)
In this study, we carried out a classification survey on sterile plants in the F1 population of high-quality and high-yield Dian-type japonica hybrid rice ‘Dianheyou 34’, combining with the basic agronomic traits and biological information of the backbone restorer line ‘Nan34’ and its mutant Rf1 restorer gene lost mutant, the phenomenon and cause of function loss of restorer lines were preliminarily explored. The results showed that the loss of function with natural mutation at Rf1 in ‘Nan34’ was the main factor that led to sterile plants (two years accounted for 0.26% and 0.79% respectively) in ‘Dianheyou 34’F1 hybrid population; the seed setting rate of ‘Nan34’ wild type was no significant difference compared with its mutant, and the wild type was slightly higher than the mutant in pollen fertility, plant height, ear length and grain length, and slightly greater than the wild type in grain width. By cloning the Rf1 locus of ‘Nan34’ wild type and its mutant, we found that the mutant in ORF region lost a 574 bp long sequence compared with the wild type, which made it unable to encode some PPR proteins. The loss of this sequence may lead to the loss of recovery function. The results lay a theoretical foundation for revealing genetic variation mechanism of restorer genes in three-line hybrid rice breeding and utilization.
Cytoplasmic male sterility (CMS) is common in some plants, characterized by aborted pollen and normal pistil development, which is inherited from the female line (Ling et al., 2000). Restoration genes in the nucleus regulate fertility restoration in plants. In the production process and cross breeding, the three-line hybrid system consisting of maintainer line, sterile line and restorer line plays a great role in the production of hybrid rice. CMS/RF system has been widely used in hybrid seed production and is helpful to elucidate the interaction between plant mitochondria and nuclear genome (Chase, 2006). There are several types of cytoplasmic male sterility, such as BT-CMS, WA-CMS, DT-CMS and HL-CMS. Genetic male sterility in rice can be divided into several types, one is the relatively common cytoplasmic male sterility (CMS), the others are photoperiod-temperature sensitive genic male sterility (PGMS), thermo-sensitive genic male sterility (TGMS) and other nuclear male sterility (Kurata et al., 2005). Previous studies have shown that proteins encoded by related nuclear genes can block cytogenic sterility genes at different levels and eliminate the effects caused by infertility genes, which are called restorative genes (Kazama and Toriyama, 2014).
Generally, restoring genes can restore fertility by changing the expression of genes related to cytoplasmic male sterility to reduce or eliminate the adverse effects caused by sterility genes (Akagi et al., 2004; Komori et al., 2004). Related studies have shown that nuclear and cytoplasmic genes jointly regulate fertility restoration of rice Dian I and BT types (Tan et al., 2004; Luo et al., 2013). It has mitochondrial chimeric gene atp6-orf79 (Kazama et al., 2008; Luan et al., 2013). The fertility of rice CMS type Dian I and BT is regulated by a nuclear restoration gene located on chromosome 10 (Akagi et al., 1996; Komori et al., 2003; Tao et al., 2004), and their sequences are 99% similar (Zhu et al., 2009). In rice (BT-CMS), the abnormal mitochondrial open reading frame orf79 is co-transcribed with a doubling of the apt6 gene, encoding the cytotoxic polypeptide ORF79. The specific accumulation of such cytotoxic peptides in microspores leads to male sterility of gametophytes (Tang et al., 2014). The Rf1 locus of rice (BT-CMS) contains two restoration genes, Rf1a and Rf1b. The proteins encoded by them are all located in mitochondria, and the generation of toxic proteins is blocked by enucleation and degradation of B-atp6/orf79 mRNA, respectively, so as to restore fertility (Wang et al., 2006). There are many reasons for the occurrence of sterile plants in hybrid rice F1 population in production, but the problem that the natural mutation of restoring genes in restorer lines may cause the loss of restoring function has not been paid attention to.
This study was aimed at the phenomenon and problem that some sterile plants with phenotype similar to hybrid rice were found in F1 field of Dian-type japonica hybrid rice 'Dianheyou34' (H479A/Nan34). By investigating the species and number of sterile plants in F1 population of hybrid rice test field, the mutation frequency of rice restoration gene under natural conditions was investigated. Then the agronomic traits of the wild type of the restorer line 'Nan34' and its mutant (loss of recovery function) were observed, and the differences in phenotypes were analyzed. At the same time, the restoration gene Rf1 locus was cloned respectively, and the differences of the two at the molecular level and the internal causes of genetic variation were compared. It provides a new idea for improving the theory and practice of utilization of heterosis of three lines in rice.
1 Result and Analysis
1.1 Natural mutation of restorer gene in Dian type rice restorer line
Rice Rf1 gene is located on chromosome 10 of rice. In the process of hybrid rice seed breeding, the restoring ability of restorer line may be lost due to some reasons. Through a detailed investigation of the species and number of sterile plants in F1 population in Xundian production demonstration field of japonica hybrid rice 'Dianheyou 34' (Figure 1), the proportion of sterile plants caused by the loss of restoring gene function of restorer lines was calculated, and then the phenomenon of natural mutation of restoring gene was preliminary determined. A total of 824 sterile plants with phenotype similar to hybrid rice 'Dianheyou 34' and 23 sterile plants with phenotype different from hybrid rice were investigated in the designated demonstration plot. The phenotype difference between the two types of sterile plants was mainly manifested in plant height. The total number of plants on the long side and the wide side were 376 and 266 respectively in the investigated rectangular experimental demonstration plot. The total number of rice plants in the experimental plot could be calculated to be about 84 970. By comparing the number of sterile plants with phenotype similar to hybrid rice and the total number of rice plants in the whole experimental field, it can be inferred that the occurrence frequency of sterile plants caused by the loss of restoring function of restorer lines is 824/84970×100%=0.97%. In conclusion, the natural mutation of restorer lines in Dian-type rice leads to the loss of restoring gene function, which is an important factor leading to the occurrence of sterile plants in F1 population. This result is similar to the identification results of Dian-type japonica hybrid rice seed purity in 2018~2019. In the F1 population of 'Dianheyou 34' produced by different households, the annual average proportion of sterile plants with similar phenotype of hybrid was 0.26% and 0.79% respectively (Table 1).
|
Figure 1 ‘Dianheyou 34’ hybrid rice production field and the sterile plants within F1 population Note: A : ‘Dianheyou 34’ hybrid production field; B: Sterile plants within hybrid F1 population; Arrow represent sterile plants |
.png)
.png) Table 1 Planting identification of seed purity of Dian-type Japonica hybrid rice in 2018~2019 Note: 1~10: Represent different farmers |
1.2 Cloning and sequencing of the restoration gene Rf1 locus of 'Nan34' and its mutants
Sequence analysis showed that the Rf1 locus of rice restoration gene was 2 971 bp in length, and the ORF region was 2 376 bp in length. It was located on chromosome 10 of rice, encoding 791 amino acids, without intron structure. Specific primers were used to amplify the restoration gene Rf1 of 'Nan34' and its mutants. The results of 1% agarose gel electrophoresis showed that the target bands of wild type and mutant were about 2 900 bp and 2 300 bp (Figure 2).
.png) Figure 2 PCR amplification at Rf1 locus of restorer line Nan 34 and its mutant Note: M: 100 bp Plus DNA Marker (Thermo Scientific); 1~2: ‘Nan34-Rf’ Wild type;3~4: ‘Nan34-rf’ mutant |
1.3 Differences in basic agronomic characters between 'Nan34' and its mutants
Plant height, seed setting rate, pollen fertility, panicle length, grain length and grain width of 'Nan34' and its mutants planted simultaneously in a greenhouse in Kunming were investigated after harvest (Figure 3).
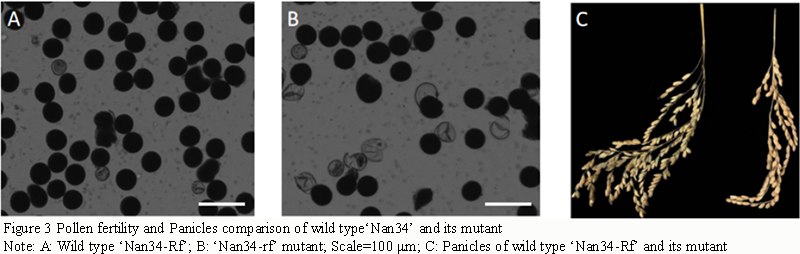 Figure 3 Pollen fertility and Panicles comparison of wild type‘Nan34’ and its mutant Note: A: Wild type ‘Nan34-Rf’; B: ‘Nan34-rf’ mutant; Scale=100 μm; C: Panicles of wild type ‘Nan34-Rf’ and its mutant |
Pollen fertility: The pollen fertility of the mutant ‘Nan34-rf’ was lower than that of the wild type 'Nan34-Rf' after I2-KI staining. The pollen fertility of the mutant 'Nan34-rf' was reduced by 5.61% compared with the wild type, and there was no significant difference in seed setting rate between the mutant and the wild type (Table 2). Plant height and panicle length (Figure 2C) of the mutant were lower than those of the wild type. The grain length of the wild type was slightly longer and the grain width was slightly shorter for the mutant, and the grain type (length-to-width ratio) of the wild type was slightly longer than that of the mutant, while the mutant was pelletized (Table 3).
.png) Table 2 Comparison of seeds rate and pollen fertility between the wild type ‘Nan34’ and its mutant Note: One-way analysis of variance, values are mean±SD; *: p<0.05 |
.png) Table 3 Comparison of basic agronomic characters of wild type ‘Nan34’and its mutant Note: One-way analysis of variance, values are mean±SD; *: p<0.05 |
1.4 Differences in the Rf1 locus of 'Nan34' and its mutants
Sequencing and comparative analysis of restorer line 'Nan34' and its mutants showed that: The amplification length of Rf1 locus of wild type restoration gene of 'Nan34' was 2 852 bp, and that of its mutant was 2 278 bp. The mutant was missing a 574 bp sequence from the ORF region of Rf1 locus of wild type restoration gene (Figure 4).
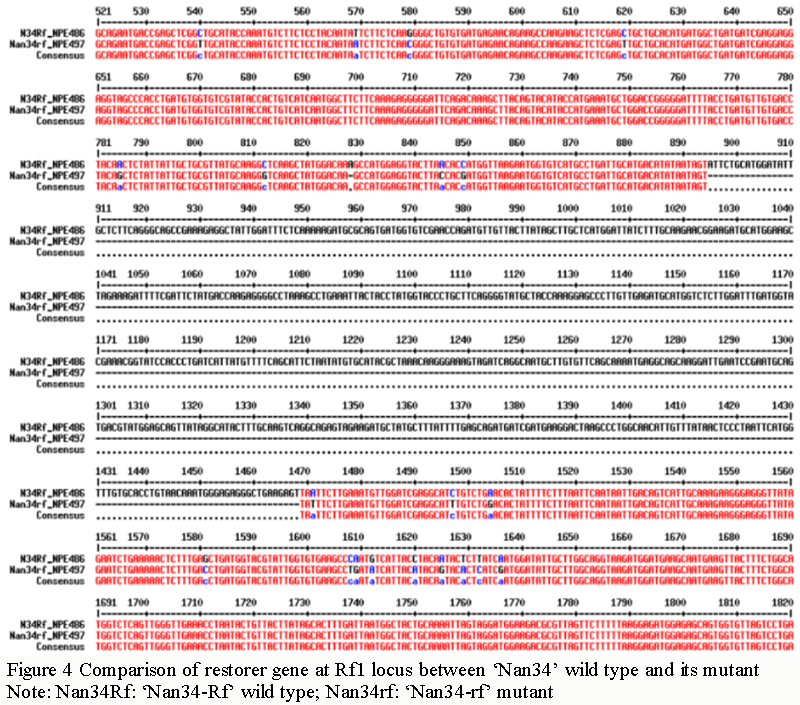 Figure 4 Comparison of restorer gene at Rf1 locus between ‘Nan34’ wild type and its mutant Note: Nan34Rf: ‘Nan34-Rf’ wild type; Nan34rf: ‘Nan34-rf’ mutant |
2 Discussion
In this study, through a detailed classification investigation of the sterile plants in the F1 population of the demonstration field production of Dian-type japonica hybrid rice 'Dianheyou34', it was confirmed that the restorer line 'Nan 34' had a natural mutation of Rf1 which resulted in the loss of the function of the restoring gene, which was the main factor leading to the occurrence of many hybrid sterile plants with similar phenotype in the hybrid F1 population. Dian-type japonica hybrid rice 'Dianheyou 34', composed of male sterile line 'H479A' and restorer line 'Nan 34', passed the variety certification in Yunnan Province in 2013. The combination has high quality and wide adaptability to disease resistance, and has been widely applied in Yunnan, Guizhou, Sichuan, Shaanxi and Wuling mountain area for many years, and is favored by farmers. However, in recent years, the phenomenon of different degrees of sterile plants was observed in the field of 'Dianheyou 34'. Related studies have found that there may be many reasons for the occurrence of population sterile plants in hybrid rice, which may be caused by human factors in the process of seed production, resulting in the similarity between sterile plants and sterile lines in phenotype (Shan et al., 2018). It may be that the restorer line fails to restore the fertility of the sterile line normally due to the loss of the restoration gene function in the process of hybridization, so the phenotype of the sterile plants is similar to that of hybrid rice (Chen and Liu, 2014). It may also be caused by photoperiod-temperature sensitive genic male sterility (PGMS) or mixed with other foreign sterile lines. However, the problem of sterile plants in hybrid rice population caused by restorer line restoration gene mutation has not been paid attention to. Through the investigation of the species and number of sterile plants in the whole experimental field of 'Dianheyou 34' planted in Xundian, Yunnan, it was found that most of the sterile plants were similar to the hybrid rice 'Dianheyou 34' in phenotype, so it was speculated that most of the sterile plants were caused by the failure of restoring gene function of restorer line 'Nan 34', which caused the infertility of the sterile line.
According to the field planting survey results of 'Dianheyou 34' in Xundian, Yunnan in 2019, it can be inferred that the occurrence frequency of sterile plants caused by the loss of restoring function of restorer line 'Nan 34' was 0.97%, indicating that the natural mutation of restoring genes has reached a certain frequency and gradually accumulated with breeding and generation. However, since the current purification cycle of restorer lines is at least 2~3 years (tasseling or paired retesting and identification in the first year, propagation in the second year, and seed production in the third year), combined with the seed purity identification results of 2018~2019 Dian-type japonica hybrid rice seeds (annual proportion of sterile plants is 0.26% and 0.79% respectively). Therefore, it can be inferred that the actual initial natural mutation frequency of the restoring gene should be at least less than 0.26%, and the initial generation and mutation frequency of the restoring gene of 'Nan 34' need to be further verified by experiments.
Compared with the wild type, plant height, panicle length and pollen fertility were decreased in mutant plants, but seed setting rate was not significantly affected. It was speculated that the mutant's restoring gene affected its growth and development through some mechanism. The full length of the wild type Rf1 locus of 'Nan 34' was 2 852 bp and the length of the ORF region was 2 376 bp. According to relevant studies, the Dian-type restoration gene and BT type Rf1a encoded 791 amino acids and were composed of 16 PPR proteins (Wang et al., 2006; Zhu et al., 2009). The mutant Rf1 locus was 2 276 bp in length, and its ORF region was missing a 574 bp sequence relative to the wild type 'Nan 34'. It was speculated that the deletion of sequence might affect the transcription and translation of some proteins, resulting in loss of recovery function. However, the frequency and mechanism of natural and tissue culture mutations in the Rf1 locus of the 'Nan 34' restorer gene need to be further studied. In the practical application of three-line hybrid rice, due to the risk of low frequency restoration gene loss in restorer line, it is suggested that the purification and retesting of restorer line should be strengthened in the process of hybrid rice seed production to reduce the risk of seed purity problems. This study provided a reference strategy for ensuring the quality of hybrid japonica rice seed production, and provided a new thinking for exploring the restoration of gene variation and molecular evolution.
3 Materials and Methods
3.1 Research materials
Restorer line 'Nan 34' and its mutant (loss of recovery function), 'Nan 34' is the core backbone restorer line of Dian-type hybrid japonica rice successfully cultivated by the Rice Research Institute of Yunnan Agricultural University in the 1990s, and its matching lines, a series of japonica hybrid rice with high quality, disease resistance, high yield and wide adaptability, such as 'Dianza 31', 'Dianza 32', 'Yuza 34', 'Dianza 35', 'Dianyou 34', 'Dianza 40', 'Dianza 80', 'Dianza 86' and 'Dianheyou 34', were promoted and applied in production. The mutant of restorer line 'Nan 34' was obtained through tissue culture progeny screening in our laboratory. T4 ligase and DNA Marker, gene cloning vector yT&A, competent Escherichia coli.
3.2 Investigation on natural mutation of restoring gene in Yunnan restorer line
The combination of male sterile line 'H479A' and restorer line 'Nan 34' resulted in a hybrid japonica rice 'Dianheyou 34', which passed the variety certification of Yunnan Province in 2013. The combination has high quality and wide resistance and adaptability, and has been popularized and applied in Yunnan, Guizhou, Sichuan, Shaanxi and Wuling mountain area for many years, and is favored by farmers. The number and species of sterile plants were observed in the experimental plots of the selected hybrid rice 'Dianheyou 34'.
Sterile plant analysis: The plants with natural seed setting rate below 10% in F1 population were determined as sterile plants. They were divided into different types according to their phenotypes: (1) The phenotype of sterile plants was similar to the investigated hybrid F1. (2) The phenotype of the sterile plant is similar to that of the female sterile line. (3) Phenotype of sterile plants was similar to heterozygous plants. The proportion of sterile plants in the total population investigated was calculated according to the types of sterile plants.
Investigation of the total population of hybrid F1: The hybrid F1 was planted in the production demonstration field of 'Dianheyou 34' in Xundian, Yunnan in 2019 (about 3 200 m2). The number of long-edge rice plants and wide-edge rice plants in the rectangular field were calculated, and the total population of rice plants in the field was roughly calculated.
By comparing the number of sterile plants with phenotype similar to hybrid rice and the total number of rice plants in the whole experimental field, the occurrence frequency of sterile plants caused by the loss of restoring function of restorer lines could be calculated.
3.3 Hybrid purity identification
Dian-type japonica hybrid rice was planted in late rice after Jingdong early rice seed production from 2018 to 2019. According to the national seed management requirements and standards, organized experts on-site identification. The seed purity identification results of Dian-japonica hybrid rice were provided by Yunnan state capital Jinding Hepu Agricultural Science and Technology Co., LTD.
3.4 Comparison of basic agronomic traits between wild type and mutant of 'Nan 34'
Pollen fertility: Three spikelets were randomly selected from wild type 'Nan 34' and its mutant before flowering. The iodine-potassium iodide staining method was used for pollen staining. Three fields were selected for each spikelet, and the number of fertile pollen and the total number of pollen were counted respectively. The percentage was calculated, and the average value was taken, and the standard deviation was calculated. Pollen fertility = Number of fertile pollen/total pollen *100%.
Seed setting rate: The main panicle of wild type 'Nan 34' and its mutant mature plants were randomly selected, the number of actual grains and total grains were counted, the average value of seed setting rate was calculated, and the standard deviation was calculated. Seed setting rate = number of solid grains/total grains *100%.
Comparison of spike length and plant height: rice pilings and main spikes of wild type and mutant mature plants of 'Nan 34' were randomly selected, respectively, and the length of each rice pilings and spikes was measured to take the average value, and the standard deviation was calculated.
3.5 Cloning of the restoration gene of wild type and its mutant of 'Nan 34'
Since the known BT type Rf1 locus in rice has no intron (Wang et al., 2006), the restoration locus was directly cloned from the nuclear genomic DNA of the mutant 'Nan34-Rf' and the wild type 'Nan 34-rf'. The full-length sequence of Rf1 locus was queried on GeneBank, and the primers Rf1Aa-F and Rf1Aa-R were designed using Primer designing tool. This gene was located on chromosome 10. DNA of wild type and mutant of 'Nan 34' was extracted by modified CTAB method. Genomic DNA was used as template, PCR amplification and electrophoresis detection were performed, and the correct target fragment was connected to the clone vector by T4 DNA ligase, and transformed into competent E. coli. After plasmid extraction and enzyme digestion, the samples were sent to BGI for sequencing (Table 4).
.png) |
3.6 Sequence alignment and analysis of restoration gene Rf1 loci of wild type 'Nan 34' and its mutant
The correct sequencing result sequence was imported into (http://multalin.toulouse.inra.fr/multalin/multalin.html) for analysis, the clonedg restorer genes loci sequence was analyzed to find out the similarities and differences of the restorer gene Rf1 locus sequence of 'Nan34' wild type and its mutants, and to analyze its molecular genetic basis.
Authors’ contributions
FT and ZQ were the executors of the experimental design and research of this study. GXQ LW and YWL completed the data analysis and the writing of the first draft of the paper. LSC, WWB, WZW, and HDJ participated in the design and analysis of the experiment results. CLJ and LXD were the originators and principals of the whole project, guiding the design of the experiment, data analysis, as well as the writing and modification of the paper. All authors read and approved the final manuscript.
Acknowledgements
This study was supported by the National Key Research and Development Program of China (2017YFD0100205), the National Natural Science Foundation of China (31560115, 31860108), the Major Project of Yunnan Province (2018ZG005) and the 2017 Yunling Talent Program - "Yunling High-end Foreign Experts" Project.
Akagi H., Yokozeki Y., Inagaki A., Nakamura A., and Fujimura T., 1996, A co-dominant DNA marker closely linked to the rice nuclear restorer gene Rf-1, identified with inter-SSR fingerprinting, Genome., 39(6): 1205-1209
Akagi H., Nakamura A., Yokozeki-Misono Y., Inagaki A., Takahashi H., Mori K., and Fujimura T., 2004, Positional cloning of the rice Rf-1 gene, a restorer of BT-type cytoplasmic male sterility that encodes a mitochondria-targeting PPRprotein, Theor. Appl. Genet., 108(8): 1449-1457
Chase C.D., 2007, Cytoplasmic male sterility: a window to the world of plant mitochondrial–nuclear interactions, Trends. Genet., 23(2): 81-90
Chen L.T., and Liu Y.G., 2014, Male sterility and fertility restoration in crops, Annu. Rev. Plant Biol., 65: 579-606
Kazama T., and Toriyama K., 2014, A fertility restorer gene, Rf4, widely used for hybrid rice breeding encodes a pentatricopeptide repeat protein, Rice, 7(1): 28
Kazama T., Nakamura T., Watanabe M., Sugita M., and Toriyama K., 2008, Suppression mechanism of mitochondrial ORF79 accumulation by Rf1 protein in BT-type cytoplasmic male sterile rice, Plant J., 55(4): 619-628
Komori T., Yamamoto T., Takemori N., Kashihara M., Matsushima H., and Nitta N., 2003, Fine genetic mapping of the nuclear gene, Rf-1, that restores the BT-type cytoplasmic male sterility in rice (Oryza sativa L.) by PCR based markers, Euphytica., 129(2): 241-247
Komori T., Ohta S., Murai N., Takakura Y., Kuraya Y., Suzuki S., Hiei Y., Imaseki H., and Nitta N., 2004, Map-based cloning of a fertility restorer gene, Rf-1, in rice (Oryza sativa L.), Plant J., 37: 315-325
Kurata N., Miyoshi K., Nonomura K.I.,Yamazaki Y., and Yukihiro I., 2005, Rice mutants and genes related to organ development, morphogenesis and physiological traits, Plant Cell Physiol., 46(1): 48-62
Luo D., Xu H., Liu Z., Guo J., Li H.Y., Chen L.T., Fang C., Zhang Q.Y., Bai M., Yao N., Wu H., Wu H., Ji C.H., Zheng H.Q., Chen Y.L., Ye S., Li X.Y., Zhao X.C., Li R.Q., and Liu Y.G., 2013, A detrimental mitochondrial-nuclear interaction causes cytoplasmic male sterility in rice, Nat. Genet., 45(5): 573–577
Ling X.Y., Zhou P.J., and Zhu Y.G., 2000, Progress of the studies on molecular machanism of cytoplasmic male sterilityin, Zhiwuxue Tongbao (Chinese Bulletin Botany), 17(4): 319-332
Luan J., Liu T.R., Luo W.Q., Liu W., Peng M.Q., Li W.J., Dai X.J., Liang M.Z., and Chen L.B., 2013, Mitochondrial DNA genetic polymorphism in thirteen rice cytoplasmic male sterile lines, Plant Cell Rep., 32(4): 545-554
Shan J.W., Cai Z.Q., Zhang Y., Xu H.N., Rao J.L., Fan Y.R., and Yang J.Y., 2018, The underlying pathway involved in inter-subspecific hybrid male sterility in rice, Genomics, 111(6): 1447-1455
Tan X.L., Tan Y.L., Zhao Y.H., Zhang X.M., Hong R.K., Jin S.L., Liu X.R., and Huang D.J., 2004, Identification of the Rf gene conferring fertility restoration of the CMS Dian-type 1 in rice by using simple sequence repeat markers and advanced inbred lines of restorer and maintainer, Plant Breeding, 123(4): 338-341
Tao D.Y., Xu P., Li J., Hu F.Y., Yang Y.Q., Zhou J.W., Tan X.L., and Jones M.P., 2004, Inheritance and mapping of male sterility restoration gene in upland japonica restorer lines, Euphytica, 138(3): 247-254
Tang H.W., Luo D.P., Zhou D.G., Zhang Q.Y., Tian D.S., Zheng X.M., Chen L.T., and Liu Y.G., 2014, The rice restorer Rf4 for wild-abortive cytoplasmic male sterility encodes a mitochondrial-localized PPR protein that functions in reduction of WA352 transcripts, Mol. Plant., 7(9): 1497-1500
Wang Z.H., Zou Y.J., Li X.Y., Zhang Q.Y., Chen L.T., Wu H.,Su D.H., Chen Y.L., Guo J.X., Luo D., Long Y.M., Zhong Y., and Liu Y.G., 2006, Cytoplasmic male sterility of rice with boro Ⅱ cytoplasm is caused by a cytotoxic peptide and is restored by two related PPR motif genes via distinct modes of mRNA silencing, The Plant Cell, 18: 676-687
Zhu J.R., Li D.X., Qian S.Q., Li J., Gan S.X., Zhou J., Qiao D.H., and Chen L.J., 2009, Cloning and genetic characterization of the fertility restorer gene Rf-D1(t) for Dian-Type cytoplasmic male sterility of rice (Oryza sativa L.), Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 7(4): 671-678



. PDF(1649KB)
. HTML
Associated material
. Readers' comments
Other articles by authors
. Tian Feng
. Qian Zhu
. Xiaoqiong Guo
. Wei Li
. Weilin Yu
. Shichuan Li
. Wenbin Wu
. Ziwei Wan
. Dajun Huang
. Lijuan Chen
. Dongsun Lee
Related articles
. Rice( Oryza sativa L)
. Dian-type CMS
. Restorer gene Rf1
. ‘Nan34’ mutant
. PPR protein
Tools
. Email to a friend
. Post a comment