Agrobacterium-mediated Transformation of Cotton Shoot Apex with SNC1 Gene and Resistance to Cotton Fusarium Wilt in T1 Generation 
 Author
Author  Correspondence author
Correspondence author
Cotton Genomics and Genetics, 2012, Vol. 3, No. 1 doi: 10.5376/cgg.2012.03.0001
Received: 18 Jan., 2012 Accepted: 02 Jul., 2012 Published: 09 Jul., 2012
Lei et al., 2011, Agrobacterium-mediated Transformation of Cotton Shoot Apex with SNC1 Gene and Resistance to Cotton Fusarium Wilt in T1 Generation, Cotton Genomics and Genetics, Vol.3 No.1 1-7 (doi:10.5376/cgg.2012.03.0001)
Using shoot apexes from the upland cotton varieties, named Zhong 35 and Junmian No.1 as explant, the SNC1, suppressor of npr1-1constitutive cloned from Arabidopsis thaliana, was introduced into cotton genome by Agrobacterium-mediated transformation with the optimized transformation conditions including culture stage of the explant, infection time and co-culture time. The results showed that the transformation procedure for high transformation rate was as follows, in vitro cuture the explants one day, infecting 20 min and co-culturing two days. PCR and RT-PCR identification of T0 and T1 regeneration plants revealed that the SNC1 gene was successfully integrated into cotton genome and expressed in the transgenic plant. Furthermore, comparing with non-transgenic plants, the T1 generation transgenic plants acquired the significant resistance to the inoculation of with the Fusarium oxysporum f.sp.Vasinfectum by the validation of the root-dipping method, which suggested that the SNC1 should facilitate the resistant to cotton Fusarium wilt.
Cotton Fusarium wilt is a destructive wilt disease in cotton production. The right way controlling cotton Fusarium wilt is to utilize the resistant germplasms and to breed resistant cotton varieties. However, it is not yet clear about the genetic regulations of cotton disease-resistant varieties, and there was a dearth of materials for resistance to Fusarium wilt in cotton germplasms. Therefore, it is difficult to breed cotton varieties resistant to Fusarium wilt through conventional breeding methods. In order to solve the problem that could not be doned with the conventional breeding methods, to expore new cotton breeding way, to breed new cotton germplasms for adapting economy development requirement, the best method is to directly introduce the required disease resistant gene into cotton genome to enhance the resistance to Furarium wilt.
Since the advent of the first case of genetically modified crops, crop genetic transformation technology has been flourishing. The application of genetic transformation technology in Cotton, the proper fiber crops, has arosen increasing attention. So far, compared with other methods of gene transformation, the Agrobacterium-mediated approach has many advantages, such as mature procedure, simple manipulation, clear transformation mechanism, high efficiency, etc. Transgenic methods can be not only to improve crop in accordance with the wishes of the people, but also to shorten the breeding periods 3 or 4 years than that of the conventional hybrid breeding methods. Up to now, Gastrodia antifungal protein gene, tobacco beta-1, 3 glucan gene and bean chitinase gene and so on have been successfully introduced into the cotton by using transgenic pathway, which have developed transgenic cotton lines with resistance to Verticillium wilt (Wu et al., 2004; Wang et al., 2004; Cui et al., 2002; Le et al., 2002; Tsai et al., 2000). It is no doubt that the genetic engineering exhibits its broad prospects for the disease resistant improvement in cotton breeding program.
At present, there are more studies on the system acquired resistance (SAR) in model plant Arabidopsis. Once plants are infected by the pathogen, the local hypersensitive resistance (HR) will generate some signaling molecules, which can induce the expression of plant defense genes. Thereby plants acquire resistance to more pathogens (Wang et al., 2004).
SNC1 (suppressor of npr1-1, constitution 1) gene cloned from Arabidopsis plays an important role in the occurrence of the plant system acquired resistance (SAR), which is negative regulator based on salicylic acid signaling transconduction, and constitutively expresses high level of PR gene with broad-spectrum disease resistance, and encodes the NB-LRR (nucletide-binding- leucine-rich repeats) protein (Li et al., 2001; Zhang et al., 2003). The NCBI Blast presents that a very little part sequence of the SNC1 gene is similar with the known cotton genome sequence, but their functions are different. The SNC1 gene is not yet to be transformed in the other plant.
Therefore, we attempted to transform the SNC1 gene into cotton through Agrobacterium-mediated using shoot apexes as explant. After obtained the transgenic cotton, Fusarium wilt strain (Fusarium oxysporum f.sp. Vasinfectum) is inoculate into the transgenic cotton to confirm whether the transgenic cotton acquire the resistance to Fusarium wilt or not.
1 Results and Analysis
1.1 The effects of explant culture time on the shoot shootapexes growth status
Picking up sterile shoot apexes cultured 0 day, 1 day, 2 days and 3 days .After infecting agrobacterium, we calculated the regeneration rates of that were from explants to green seedlings. shoot apexes were shorter , only 3~4 mm in length, so they were caused seriously damage after maken traumas, The statistical results showed that the regeneration rate of shoot apexes of Zhong 35 and Junmian No.1 cultured 0 day were 44.1% and 43.7%, respectively; While cultured 2~3 days, shoot apexes began to grow slowly, and were difficult to recover after maken traumas, its generation rate were 54.2% to 70.0%, respectively; whereas shoot apexes became plump and 6-8mm in length, and grew rapidly afer maken traumas , and the generation rate of shoot apexes of Zhong 35 and Junmian No 1 cultured in one day reached 80.9% and 81.5%, respectively.(Table 1).
Table 1 Effect of explant-cultured time on the growth status of shoot apex |
1.2 The effect of bacterium infection time on the shoot apex growth status
Using Dissecting needle tip to make traumas on meristem tissue of shoot apexes, agrobacterium infection time were set 10 min, 20 min and 30 min. We found that the generation rate of sterile stem apexes of Zhong35 and Junmian No.1 infected by 10 min were 56.1% and 62.0%, respectively, and infected by 30 min the explants grew up slowly, the seedlings were small and dwarf; whereas the generation rate of sterile stem apexes of Zhong35 and Junmian No.1 infected by 20 min reached 76.7% and 74.6%, respectively (Table 2).
Table 2 Effect of infection time on the growth status of shoot apex |
1.3 The effect of co-cultur time on shoot apex growth status
The co-culture time was set 1 day, 2 days, 3 days and 4 days . The results showed that the morphology of all shoot apexes unchanged after 1 day, and the top of shoot apexes became expanding and thickening; cultured for 3 days and 4 days, Apical elongation was up to 20~30 mm , and the middle of the apexes dehydrated and were severe browning; cultured for 2 days shoot apexes grew up about 10 mm, after inoculated on the selective medium, the top of apexes gradually became light yellow, it was obvious that the restoring time for their growth was short and young buds were robust, the regeneration rate of apexes of Zhong 35 and Junmian No.1 can reach 66.2% and 71.9% , respectively(Table 3).
Table 3 Effects of co-culture time on the growth status of shoot apex |
1.4 Obtained transgenic SNC1 cotton
The transgenic cotton seedlings being rooting were planted in the flower pots that filled with the steriled potting loam and vermiculite (mixed by 1:1) and irrigated nutrient solution prior to bagging, and placed in the incubator to continuing foster. After 10 days, healthy cotton seedlings were transplanted into the greenhouse (Figure 1). The results showed that the 45 individuals were survived, and the 16 individuals were proved to be transgenic SNC1 cotton plants by PCR assays (Figure 2). Transgenic cotton seeds were harvested respectively according to the different varieties.
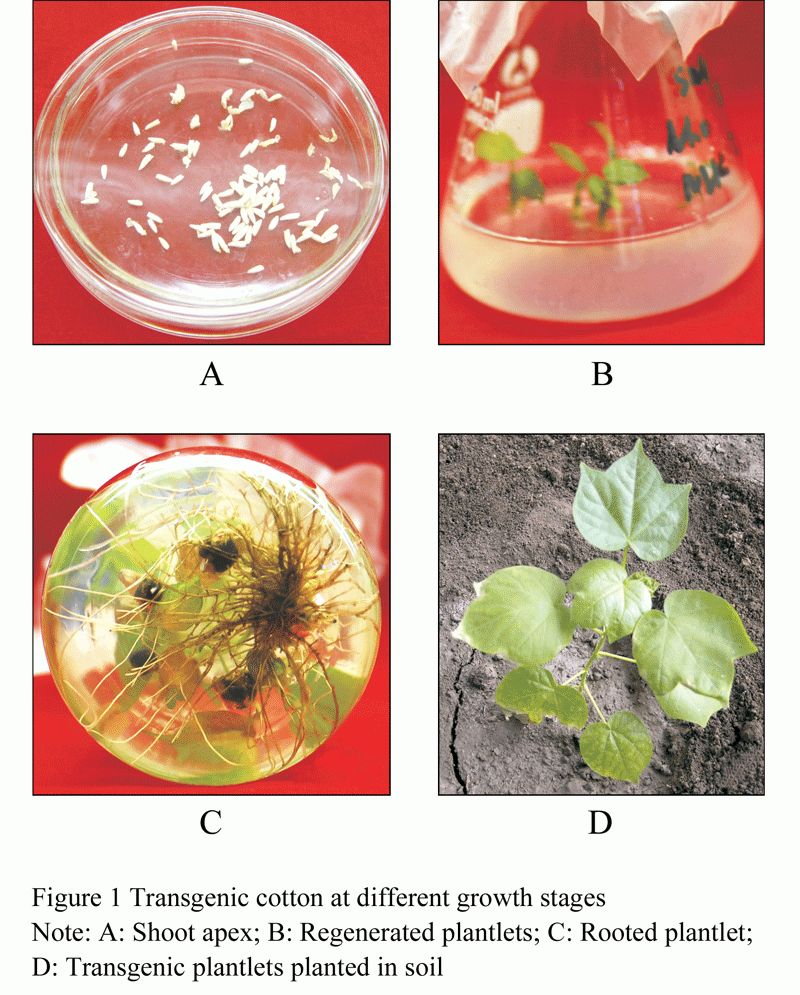 Figure 1 Transgenic cotton at different growth stages |
Figure 2 Detection of SNC1 gene in part of the transgenic plants by PCR |
1.5 PCR detection of T1 generation of transgenic SNC1 cotton
All T0 generation seeds of transformed plants were planted , when the cotton grew up in the five-leaf stage, genomic DNA of T1 generation cotton was extracted form leaves, PCR detection for the target gene and selectable marker Bar gene were carried out.
The PCR product of the target gene SNC1 by the electrophoresis showed that 281 of 860 plants had a single clear band with 790 bp that located in the same position as the plasmid amplification products; while there was no band appearing in the non-transgenic cotton. There were 153 individuals in 281 transgenic plants belonging to transgenic Zhong 35 , the rest 128 individuals were transgenic Juanmian No.1. It might be proved that the target gene had been integrated into the genome of the cotton plant with a definit genetic stability.
The PCR product of the marker Bar gene was amplified. The electrophoresis results showed that transgenic plants had a clear band with 146 bp that located in the same position as the plasmid amplification products. There were 153 individuals of Zhong 35 and 128 individuals of Juamin No.1, whereas no band amplified in non-transgenic plants (Figure 3). It is obvious that the PCR detection result of marker Bar gene was consistent with that of SNC1 gene.
Figure 3 Detection of selective marker Bar in part of the transgenic plants by PCR |
1.6 RT-PCR detection of T1 generation of transgenic SNC1cotton
When the transgenic cottons grew up with the six-leaf stage, total RNAs of 281 individuals of the T1 generation with SNC1 gene were extracted from leaves for RT-PCR detection. The target gene SNC1 with the band of 790 bp at the transgenic plants were detected with the same size as the positive plasmid, which indicated that the SNC1 gene in the T1 generation had expressed in the RNA level (Figure 4).
Figure 4 Detection of some SNC1 transgenic plants by RT-PCR |
1.7 Identification of disease resistance for T1 generation of transgenic SNC1 cotton
A certain concentration of cotton Fusarium wilt strain (Fusarium oxysporum f.sp. Vasinfectum) was inoculated in the cotton seedling by root-dipping. The rates of disease incidence were recorded every 5 days until the incidence of disease has been stable in the 21 days. The rate of incidence showed that in the 21days, the incidence rate of Juanmian No.1 as control (non-transgenic cotton) had 66.7%, while the incidence rate of transgenic Juanmian No.1 had 37.5% that is about half of the control; Whereas the incidence rate of Zhong 35 as control (non-transgenic cotton) had 50.0%, while the incidence rate of transgenic Zhong 35 had 22.2% that is less than half of the control. This result indicated that the transgenic SNC1 cotton varieties should acquire obvious disease-resistance compared with non-transgenic cotton varieties, while transgenic Zhong35 had much more significant disease resistance than that of the transgenic Juanmian NO.1(Table 4; Figure 5).
Table 4 The incidence of Fusarium oxysporum f. sp vasinfectum in transgenic and non-transgenic plants after inoculation |
.png) Figure 5 Symptom of transgenic and non-transgenic plants after inoculation in 21days |
2 Discussion
Although the Agrobacterium-mediated method is a more mature transgenic approach, only a few in vitro culture materials of varieties can be regenerated because of long experimental period and limits of genotypes in somatic regeneration in vitro. Cotton shoot apex culture for regeneration is relatively easy due to using complete meristem. Since Renfroe et al. reported that they obtained the regenerated plants using cotton shoot meristem, many scholars at home and abroad reported the evidences of transgenic plant regeneration derived from the apical meristem (Sun et al., 2009; Zhao 2009; Weng et al., 2009; Zhou et al., 2009; Lv et al., 2004; Balasubrmani et al., 2003).
In this study, we transformed SNC1 into cotton using shoot apexes by Agrobacterium-mediated method.Through modifying the growth stage, the infection time, co-culture time, which greatly improved the efficiency of genetic transformation, and we obtained transgenic cotton.
By PCR and RT-PCR detection, we verified that the exogenous target gene has integrated into the cotton genome, and has expressed in the level of transcription.
We selected a strong virulence strain of Fusarium oxysporum f.sp. Vasinfectum in Xinjiang, which pathopoiesis was up to 66.7% to susceptible variety Junmian NO.1. Transgenic SNC1 cotton varieties were significantly lower incidence than that of non-transgenic controls, indicating that transgenic SNC1 cotton acquired cotton Fusarium wilt resistance. However, this was the indoor results of T1 generation plants, and we needed to verify resistance of transgenic cotton in field disease nursery .
3 Materials and Methods
3.1 Receptor materials
The receptor materials were the Southern of Xinjiang main cultivated upland varieties, Zhong 35 and Juanmian NO.1, which provided by the Key Laboratory of Crop Biotechnology, the Institute of Nuclear Biological Technology, Xinjiang Academy of Agricultural Sciences.
3.2 Plasmids and Agrobacterium strains
Agrobacterium strain GV3101 containing SNC1 gene was provided by Dr. Li Xin from University of British Columbia, Canada. Strain was selected with kanamycin 50 mg/L. Gentamicin 50 mg/L and rifampicin 50 mg/L, and the herbicide as selection marker used for the resistance of regeneration plants.
3.3 Reagents
Conventional reagents were domestic analytical grade. Reverse transcriptase were purchased from Promega Corporation. 10× Bufffer, dNTPs and Taq enzymes were purchased from Shanghai Biological Engineering Technology Co., Ltd..; the SNC1 primers for PCR detection were synthesized by the Beijing Huada Genomics Research Center.
3.4 Agrobacterium-mediated genetic transformation
The de-fuzz seeds were sterilized with alcohol for 30 seconds, and then with hydrogen peroxide disinfection for 2 ~ 3 hours, finally rinsing with sterile water with 3~4 times. Using one-day cultured shoot apexes for the infection for 10 min, co-cultured for 2 days. The MS basic medium with additional IAA 0.1 mg/L, KT 0.2 mg/L, as the basic culture conditions, while the optimization for shoot apexes culture, the infection time and co-culture time was carried out.
Taking the 0 day and 3 days cultured shoot apex to be infected 10~30 min (agrobacterium concentration OD600 = 0.5),dark-cultured 1 to 4 days, and then transferred into induction medium with the herbicide (0.1 percent) and Cefotaxime (500 mg/L), cultured in the lighting at 28℃. Experiments were repeated three times to record the rate of regeneration of explants that grew up green seedlings. When the shoots grew up 1~2 cm in length (about 20~30 days), the first subculture were carried out, followed by 2~3 times for continuing subcultures until the shoots grew to 3~4 cm in length (about 30~60 days), then transferred to rooting medium (about 30 ~ 40 days). The seedlings grew young radicles were transplanted for seedling recovery. The whole process from seed disinfection to regenerated plant takes 3~ 5 months.
3.5 Molecular detection of transgenic plants
Total DNA was extracted from the leaves of herbicide-resistant plants by CTAB method (Paterson, 1993). PCR analysis for herbicide resistant plants were carried out by using SNC1 gene-specific primers (designed based on the conserved region of its sequence) and herbicide marker primers. The PCR reaction system is 2.0 μL 10×Taq polymerase corresponding buffer 1.6 μLdNTPs, (2.5 mmol/L), each primer 0.3 μL, 0.2 μL Taq DNA polymerase, 1 μL DNA templates in total 20 μL reaction volume. The cycling parameters was indicated as below: 94℃ 3 min, each circle was 94℃ 30s, 51℃ 45 s (SNC1), 72℃ 1 min, 30 circles,and extend 72℃ 10 min. Basta cycling parameters with a melting temperature of 55℃ 30 s
Amplified product were scored by 1% agarose gel electrophoresis. Specific primers for SNC1 with expected fragment size about 790 bp were designed as SNC1-F: 5'-CCAAACATTTTCAGACTTACAAGACTTG-3 ' and SNC1-R: 5'-AGATGTCCCCGATGTCATCC-3'; for herbicide primers with expected fragment size 146 bp as Basta-F: 5'-AGCCCGATGACAGCGACCAC-3 ', and Basta-R: 5'-CGCAACGCCTACGACTGGAC-3'.
3.6 T1 generation of transgenic plants identified by RT-PCR
Total RNA was extracted from the cotton leaves of T1 generation plant identified by PCR (Liu et al., 2006), the SNC1 gene were detected by RT-PCR using Promega's reverse transcription system.
3.7 Disease resistance of transgenic plants identified by inoculation
Root-dipping was followed the description of Plant Pathology Research Methods composed by Fang Zhongda. Mixes vermiculite and potting loam inaccording to the ratio (1:1) , T0 generation of transgenic Zhong 35 and transgenic Juanmian No.1 and their corresponding non-transgenic cotton were sowed in the nutrition pots, respectively. When the T1 generation transgenic cottons and non-transgenic cotton grew up 3-leaf stage, after injuried root, inoculated with Fusarium oxysporum strains (provided by Professor Li Guoying from Shihezi University), of which spore suspension was diluted to 1.0×106 /mL. The incidence of transgenic and non-transgenic cotton was begun to record every five day until 21days, and to score the number of the incidence and to calculate the incidence rate.
Acknowledgements
We are very grateful to Dr. Li Xin from University of British Columbia, Canada, kindly presented SNC1 gene used in this study. This project was funded by the Transgenic Special Project (2009ZX08005-011B) of the Ministry of Agriculture of China.
References
Balasubrmani G., Amudha J., Kumar P.A., and Mayee C.D., 2003, Agrobacterium-mediated transformation in indian cotton (G. hirsuturn) cultivar with crylA(b) gene and regeneration by direct shoot organogenesis, Mianhua Xuebao (Cotton Science), 15(1): 51-58
Cui B.M., Zhu J.B., Xiao L., Wang A.Y., Zhu X.X., Le J.H., and Chen Z.H., 2002, Cotton (Gossypium hirsutum) transformed with β-1,3-glucanase gene and chitinase gene, Shengwu Jishu (Biotechnology), 12(4): 112
Cai Y.F., Ye P.S., Jiang H.Z., He H.H., Tan Y.J., Pei Y., Guo Y.L., Li M.Y., and Hou L., 2000, The preliminary study of anti-fungi genes introduced to cotton to develop transgenic cotton with high resistance to Verticillium wilt, Xi'nan Nongye Xuebao (Southwest China Journal of Agricultural Sciences), 13(4): 45-49
Le J.H., Zhu J.B., Cui B.M., Zhu X.X., Wang A.Y., Chen Z.H., Liu G.Z., Li G.Y., Jian G.L., and Wu G., 2002, Breeding fusarium wilt and Verticillium wilt of cotton-resistant strains with transgene technique, Shihezi Daxue Xuebao (Journal of Shihezi University (Natural Sciences)), 6(3): 173-178
Li, X., Clarke, J.D., Zhang, Y., and Dong X., 2001, Activation of an EDS1-mediated R-gene pathway in the snc1 mutant leads to constitutive, NPR1-independent pathogen resistance., Molecular Plant-Microbe Interaction, 14: 1131-1139
http://dx.doi.org/10.1094/ MPMI.2001.14.10.1131 PMid:11605952
Liu Y., He X.R., Ma H.B., Wu Y.L., and Yang Y.M., 2006, Extraction of total RNA from cotton (Gossypium hirsutum L.) tissues with CTAB-PVP method, Zhongguo Nongye Daxue Xuebao (Journal of China Agriculture University), 1(1): 53-56
Lv S.L., Yin X.Y., Zhang K.W., and Zhang J.R., 2004, Agrobacterium-mediated transformation of shoot apex of cotton and production of transgenic plants carrying betA gene, Gaojishu Tongxun (High Technology Letters), 1: 20-25
Paterson A.H., 1993, A rapid method for extraction of cotton (Gossypium ssp.) genomic DNA suitable for RFLP or PCR analysis, Plant Mol. Biol. Rep., 11: 122-127
http://dx.doi. org/10.1007/BF02670470
Sun X., Su X., Ling L., Cai Y., Hong S.L., Lin Y., and Cai Y.P., 2009, The effect of Agrobacterium-mediated transformation system of shoot apex on transformation frequency of different genotypes of cotton, Zhongguo Nongxue Tongbao (Chinese Agricultural Science Bulletin), 25(7): 62-66
Wang Y.Q., Chen D.J., Wang D.M., Huang Q.S., Yao Z.P., Liu F.J., Wei X.W., and Li R.J., 2004, Over-expression of Gastrodia anti-fungal protein enhances Verticillium wilt resistance in coloured cotton, Plant Breeding, 123: 454-459
http://dx.doi.org/10.1111/j.1439-0523.2004. 01005.x
Wang Z.H., Jia Y.L., and Xia Y.W., 2004, Research advances on molecular mechanism of disease resistance in plant, Zhiwuxue Tongbao (Chinese Bulletin of Botany), 21(5): 521-530
Weng Q., Chen Q.J., Kong Q.P., Zhang G.Z., Wu J.L., and Qu Y.Y., 2007, Study on regeneration system and influencing factors of genetic transformation of G. barbadense shoot apex mediated by Agrobacterium, Xinjiang Nongye Daxue Xuebao (Journal of Xinjiang Agricultural University), 30(4): 63-67
Wu J.H., Zhang X.L., Luo X.L., Nie Y.C., Tian Y.C., and Chen Z.H., 2001, Transgenic cotton plants of chitinase and glucanase genes and their performance of resistance to Verticillium dahliea, Yichuan Xuebao (Acta Genetica Sinica), 31(2): 183-188
Zhang Y., Goritschnig S., Dong, X., and Li X., 2003, A gain-of-function mutation in a plant disease resistance gene leads to constitutive activation of downstream signal transduction pathways in the snc1 mutant, Plant Cell, 15: 2636-2646
http://dx.doi.org/10.1105/tpc. 015842 PMid: 14576290 PMCid:280567
Zhao F.Y., 2009, Establishment of Agrobacterium-mediated transformation system of cotton shoot tip, Anhui Nongye Kexue (Journal of Anhui Agricultural Sciences), 37(2): 515-518
Zhou X.F., Jin S.X., Li X., Li B.C., Lian W.M., Ning X.Z., and Liu L., 2009, Multiple-shoot induction and genetic transformation of island cotton (Gossypium barbadense L.) in Xinjiang, Mianhua Xuebao (Cotton Science), 21(4): 324-329



. PDF(279KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Jiangrong Lei
. Xiaorong Li
. Dongmei Wang
. Lin Shao
. Xiaowei Wei
. Leping Huang
Related articles
. Upland cotton
. SNC1
. Agrobacterium -mediated transformation
. Fusarium wilt
Tools
. Email to a friend
. Post a comment