QTL Mapping and Analysis of Traits Related to Pod Dehiscence in Soybean 
2. Alkali Soil Natural Environmental Science Center (ASNESC), Northeast Forestry University, Harbin, 150040, P.R. China
3. Industrial Crop Research Institute, Shaiaxi Academy of Agriculture Sciences, Fenyang, 032200, P.R. China
4. Hainan Key Lab of Crop Molecular Breeding, Hainan Institute of Tropical Agricultural Resources (HITAR), Sanya, 572025, P.R. China
 Author
Author  Correspondence author
Correspondence author
Legume Genomics and Genetics, 2012, Vol. 3, No. 3 doi: 10.5376/lgg.2012.03.0003
Received: 15 Jun., 2012 Accepted: 16 Jul., 2012 Published: 23 Jul., 2012
Luo et al., 2012, QTL Mapping and Analysis of Traits Related to Pod Dehiscence in Soybean, Legume Genomics and Genetics, Vol.3, No.3 14-20 (doi: 10.5376/lgg.2012.03.0003)
Pod dehiscence (Pod shattering) (PD) is an important reproductive trait in soybean, while this trait also causes major yield losses in the mechanically harvested soybeans. In this research, QTL mapping and correlation analysis of three related traits (PD trait (PDH), ratio of thickness to width (RTW) and days of full maturity (DFM)) were performeded in 112 strains of soybean recombinant inbred lines (RIL) population (JINF population) by using methods of composite interval mapping (CIM). The results showed that there was an extremely significant negative correlation between PDH and RTW (P=0.01, -0.960), a significant negative correlation between PDH and DFM (P=0.05, -0.740), and also a significant positive correlation between RTW and DFM (P=0.05, 0.867). The results of QTL mapping demonstrated that there was a major QTL controlling PD, designated as qPDH6-1, mapped on soybean molecular linkage group (LG) C2 (chromosome 6) between Sat_062 and Satt520 with a linkage distance of 3.1 cM. The LOD value was found to be 12.15, and about 49.44% of the total variance was accounted for. Total of 3 QTLs linked to DFM, designated as qDFM6-1, qDFM6-2 and, qDFM18-1, were examined by CIM. qDFM6-1 was found in the same marker region between Sat_062 and Satt520 as qPDH6-1 on LG C2 (Chromosome 6), with linkage distance of 7.2 cM , LOD value of 6.59, and 21.77% of total variance accountable. qDFM6-2 was also found on chromosome 6 in the region between Satt291 and Satt305 with linkage distance of 12.3 cM, LOD value of 2.85, and 9.74% of the total variance accounted for. qDFM18-1 was mapped on LG A2 (Chromosome 18) in the region between Satt217 and Satt130,with linkage distance of 0.6 cM, LOD value 2.81, and 7.74% of the total variance accounted for. 3 QTLs related to RTW were also identified, designated as qRTW11-1, qRTW4-1, and qRTW8-1, which were mapped in the regions between the SSR markers Satt426 and Satt509 on LG 11, between Satt361 and Satt399 on LG 4 and between Satt187 and Satt377 on LG 8, with the corresponding linkage distances of 7.2 cM, 12.3 cM and 0.6 cM, LOD values 3.00, 2.77 and 2.14, and 9.03%, 8.20% and 7.02% of the total variance accounted for, respectively.
Pod dehiscence (PD) of soybean refers to the trait of mature pods opening along the dorsal or ventral sutures to release seeds. Pod dehiscence (PD) is achieved through evolution so that soybeans can better produce offsprings, and it is still commonly observed in the wild soybean species. In order to harvest soybean seeds effectively, people eliminated the species which are easy to shatter in the process of domestication in cultivated soybean. Although to cultivate the non-shattering soybean species would be the major goal for soybean cultivar, in the case of delayed harvesting or dry weather, there are still many soybean pods shattering their seeds naturally. This will cause a lot of yield losses in soybean harvesting.
Scientists have constructed the soybean recombinant inbred lines (RILs) by using resistant and susceptible cultivar as parents, and they used the RILs to identify the QTL for the PD. Bailey et al (1997) identified 12 restriction fragment length polymorphism (RFLP) markers for PD, which were located on chromosome 2, chromosome 15, chromosome 16 and chromosome 19, respectively. Funatsuki et al (2006) mapped a QTL for PD in RILs by CIM method, designated as qPDH1, which was located between the SSR markers Sat_093 and Sat_366 on chromosome 16with genetic distance of 2.9 cM. In 2008, they confirmed that this QTL was located between Sat_093 and Sat_366 on chromosome 16 using soybeans from 4 different genetic backgounds, which accounted for 50% of the total variance. They also found that the allelic gene controlling PD has multiple-bit points in different genetic backgrounds (Funatsuki et al., 2008). Suzuki et al (2010) thought that qPDH1 controls the process of PD in soybean, while the morphological characteristics of soybean didn’t relate to this process. Yamada et al (2009) analyzed the PD among several soybean populations, and they found that there were differences among the PD controlling genes in different genetic backgrounds, but qPDH1 was still the gene responsible for controlling PD. Kang et al (2009) identified one major QTL and three minor QTLs for PD, which were located on chromosome 6, chromosome 5, chromosome14 and chromosome 10, respectively. The major QTL was located near to qPDH1. They also found that PD controlling genes have some differences in different genetic backgrounds.
In this study, we used the RILs population (JINF population) constructed by our laboratory and Shanxi Academy of Agricultural Sciences (Liu et al., 2003) to perform QTL mapping and correlation analysis on PD and 2 other related traits, namely days of full maturity (DFM) and Ratios of thickness to width (RTW) so that we can understand the relationship between PD trait and other related traits. This will help lay down the foundations for future application on gene cloning and molecular breeding.
1 Results and Analysis
1.1 Correlation analysis among the three related traits
We surveyed the phenotypic evaluations of PD, days of full maturity (DFM), ratio of thickness to width (RTW) in 112 strains of soybean JINF population, and we analyzed the correlation analysis of these three related traits with SPSS 16.0. The results demonstrated that there was a certain negative correlation between PD and the DFM and RTW, an extremely significant negative correlation between PD and RTW (P=0.01, -0.960), and a significant negative correlation between PD and DFM (P=0.05, -0.740). Nevertheless, there was a significant positive correlation between RTW and DFM (P=0.05, 0.867).
1.2 QTL mapping of PD
We identified the QTL location for the PD in 112 soybean strains of JINF population by the composite interval mapping (CIM) method in WinQTLCart 2.5. Set the LOD value>2.0. We obtained a QTL for the PD, designated as qPDH6-1, which was located in the region between Sat_062 and Satt520 on the linkage group C2 (Chromosome 6) with LOD value 12.15, genetic distance of 3.1 cM, and 49.44% of the total variance accounted for (Figure 1; Table 1).
.png) Figure 1 Location of the QTL for PD on chromosome 6 in the JINF RIL population |
Table 1 The information about QTLs of PD, RTW and DFM in the JINF RIL population |
1.3 QTL mapping of RTW
Three QTLs location were identified for the RTW in 112 soybean strains of JINF population by the composite interval mapping (CIM) method in WinQTLCart 2.5, which were designated as qRTW11-1, qRTW4-1 and qRTW8-1, respectively (Figure 2; Table 1).
Figure 2 Location of three QTLs for the ratio of thickness to width on chromosome 11, 4 and 8 in the JINF RIL population |
qRTW11-1 was located in the region between Satt426 and Satt509 on the linkage group B1 (chromosome 11) with LOD value 3.00, genetic distance of 10.0 cM, and 9.03% of the total variance accounted for. qRTW4-1 was located in the region between Satt361 and Satt399 on the LG C1 (chromosome 4) with LOD value 2.77 and genetic distance of 0.4 cM, and it could account for 8.20% of the total variance. qRTW8-1 was located in the region between satt187 and Satt377 on the LG A2 (chromosome 8) with LOD value 2.14 and genetic distance of 3.7 cM, and it accounted for 7.02% of the total variance.
1.4 QTL mapping of DFM
We also identified the QTL location for DFM in 112 soybean strains of JINF population by the composite interval mapping (CIM) method in WinQTLCart 2.5. Setting The LOD value>2.0, we acquired three QTLs for DFM, which were designated as qDFM6-1, qDFM6-2 and qDFM18-1 (Figure 3; Table 1).
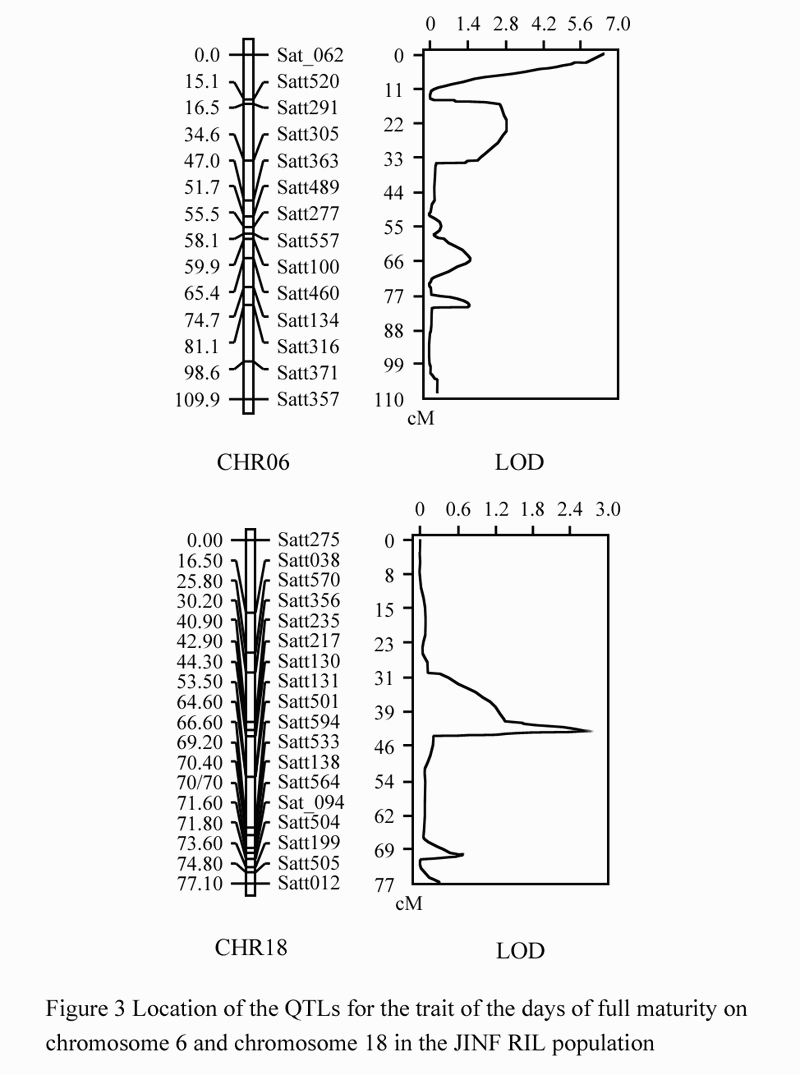 Figure 3 Location of the QTLs for the trait of the days of full maturity on chromosome 6 and chromosome 18 in the JINF RIL population |
Both qDFM6-1 and qDFM6-2 were located on the linkage group C2 (chromosomes 6). They were located in the region between Sat_062 and Satt520, and between Satt291 and Satt305, with the LOD values 6.59 and 2.85, the genetic distances of 7.2 cM and 12.3 cM, and accounted for 21.77% and 9.74% of the variance, respectively. qDFM18-1 was located in the region between markers Satt217 and Satt130 on the linkage group G (chromosome 18) with the LOD value 2.81, the genetic distance of 0.6 cM, and accounted for 7.74% of the variance.
1.5 Analysis of genomic information of related QTLs
Based on the Genome ANNOTATION INFORMATION of soybean on http://www.phytozome.net/cgi-bin/gbrowse/soybean/, we found a gene that have the auxin response protein activity, Glyma06g00860.1, and a gene encoding hydrolytic enzyme protein, Glyma06g01500.1, near the location of qPDH6-1. We also found a gene encoding auxin transfer protein, Glyma04g43150.1, and another gene encoding endo-polygalacturonic acid enzyme, Glyma04g43340.1, near the location of qRTW4-1. There was also an endo-polygalacturonic acid enzyme gene, Glyma18g16870.1, near the location of qDFM18-1.
2 Discussion
Most previous studies suggested that the major QTL (qPDH1) for PD in soybean was mapped on soybean molecular linkage group J (chromosome 16) (Funatsuki et al., 2006; 2008; Liu et al., 2007; Kang et al., 2009; Suzuki et al., 2010). Kang et al (2009) reported that different breeding strategies will be required for PD depending on different genetic backgrounds (Kang et al., 2009). Our present research result showed that there was discrepancy from previous studies in QTL mapping, and the major QTL (qPDH6-1) was mapped on chromosome 6. This is probably due to the testing materials in the study, RIL population derived from across between ‘Huibuzi’ (Glycine max Merr., CV. ZDD2315)בJidou 23’ (Glycine max Merr., CV. Jinbean 23) and a pod shattering susceptible cultivar ‘Huibuzi’.
In present mapping study, we found more than one QTLs located on soybean molecular linkage group C2 (chromosome 6), and a major QTL for PD (qPDH6-1) located on Chr. 6 with the genetic distances ranging from 0.4 cM to 3.6 cM, a major QTL for DEM (qTFM6-1) located from 0.1 cM to 7.3 cM on Chr. 6. Both QTLs were located in the region between Satt_062 and Satt520. In addition, a major QTL for DFM (qDFM6-1) was located on Chr. 6 with the genetic distance from 16.4 cM to 28.7 cM. The above 3 QTLs showed overlapping or adjacent position indicating that there might be many candidate genes controlling PD located on the Chr.6 in soybean.
Tsuchiya (1987) reported that the ratio of width to thickness of the resistant varieties was higher than those of the susceptible varieties, and the size and shape of pods may not directly related to PD. Caviness’s (1969) study showed that the value of width/thickness and PD were related to seed size. In this study, there was a significantly negative correlation among PD, the value of width/thickness and date of maturity which confirmed the results from previous researches.
Arnaud and his colleague’s study showed that synthesis of the phytohormone gibberellin has a key role in patterning the Arabidopsis fruit and it is required for fruit opening (Arnaud et al., 2010). Based on this result and analysis of bioinformatics of mapped QTLs, we found a gene Glyma06g00860.1 with protein activity in gibberellin response located on Chr. 6 near the qPDH6-1, and a gibberellin transfer protein gene Glyma04g43150.1 located on Chr. 4 near the qRTW4-1. Although it was unclear what the functional annotations of these two predicted genes were and what their protein products were, we suggested that gibberellins and its relative synthesis genes might play a key role in PD process of soybean.
Christiansen et al (2002) found that the genes encoding for the polygalacturonase (PG) were different between soybean and rapeseed, but Petersen et al., (1996) demonstrated that the polygalacturonase was related to PD in rapeseed. Coincidentally, we found two polygalacturonase genes encoding for activity protein, which were Glyma04g43340.1 l near the qRTW4-1 on chromosome 4 and Glyma18g16870.1 near the qDFM18-1 on chromosome 18, respectively. This result implied that PG might play a role in the PD process of soybean, and it should be a new direction for further research.
Scientists thought that hydrolase plays a key role in the process of PD (Fry et al., 1992; Agrawal et al., 2002), and we found a gene Glyma06g01500.1 encoding for the hydrolytic enzymes located on Chr. 6 near to the location of qPDH16-1.
Although the functional annotation of above predicted genes, and whether their protein products related to PD remained unclear, we predicted that gibberellins, relevant PG and hydrolytic enzymes might play a different role in PD regulatory network in plant. In addition, Arnaud et al (2011) showed that it would be helpful to regulate fruit opening and seed dispersal by changing the gene sequence of promoters, which would provide an important study direction and material for further research in PD relative genes and their regulatory network.
3 Materials and Methods
3.1 Materials used in this research
The soybean female parent was Glycine max Merr., The CV Jinbean 23 which does not shatter easily after maturation; the male parent was pod shattering susceptible cultivar, Glycine max Merr. CV. ZDD2315, which shatters easily after maturation. The recombinant inbred lines (JINF population) were constructed from the female and male parents. All of these materials were provided by Xueyi Liu from Industrial Crop Research Institute, Shanxi Academy of Agriculture Sciences.
3.2 Genetic map of soybean and QTL analysis
Wan et al (2005) used the JINF population as materials, and integrated a soybean genetic linkage map which has 227 SSR markers. Based on the SSR markers in this map and the markers in soybean public map, Gong et al (2006) re-integrated a new genetic linkage map. In this research, 240 SSR markers from the soybean molecular genetic map constructed by Gong (2006) were used to identify the QTLs for the related traits.
Open the "Frequencies" dialog box in "Descriptive Statistics" under "Analyze" with SPSS 16.0, select"Histogram" and "With normal curve", and make the column charts for the three related traits. Next, open=the "Bivariate" in "Correlate" under "Analyze" with SPSS 16.0, select the "Means and standard deviations" and "Excludes cases pairwise" in "Options" box, set the "Correlation Coeffecients" as "Pearson", select "Two-tailed" in "Test of Significance", and analyze the correlations of the three related traits. Import the data to the WinQTLCat 2.5 software (http://statgen.ncsu.edu/qtlcart/WQTLCart.htm), set the "Walk speed" as 2, and "By Manual Input" as 11.5, the rest boxes as default values to map the QTLs for the three related traits by CIM.
3.3 Soybean planted in the field
The materials for the research were dried in the sun for one day. The seeds were planted at the square pots (20 cm×20 cm) of Hainan Institute of Tropical Agricultural Resources (HITAR), on August 5th, 2011. Each pot has 4 kg clay (The clay also needs to be dried in the sun for one day and mixed with fertilizer). Three pots for each parent, and the one pot for each of the 112 strains were grown. 5~6 seeds were planted in each pot, and final singling was performed when seedlings reach V4 stage. Three strains were kept in each pot. During seedling growth, watering and weeding of the plants were performed periodically, and the data record and harvest were completed at relevant growing stages.
3.4 Phenotype dermination
3.4.1 Phenotype assessment of PD
The pods were collected (10~20 pods from each plant) when the soybean was grown to R8 stage (95% of the pods have reached their mature, brown or tan color), and all the pods were dried in room for 15 days.
The degree of PDH was evaluated by monitoring the percentage of the dehiscent pods after heat treatment at 80℃ for 5 hours (Tukamuhabwa et al., 2002). In the process of heat treatment, in order to prevent the pods burst affect the outcome statistics, pods were put into a flask first. The percentage of shattering pods was then recorded, and average was obtained from the 3 replicates of each strain. The cylindrical map was created with SPSS 16.0 (Figure 4).
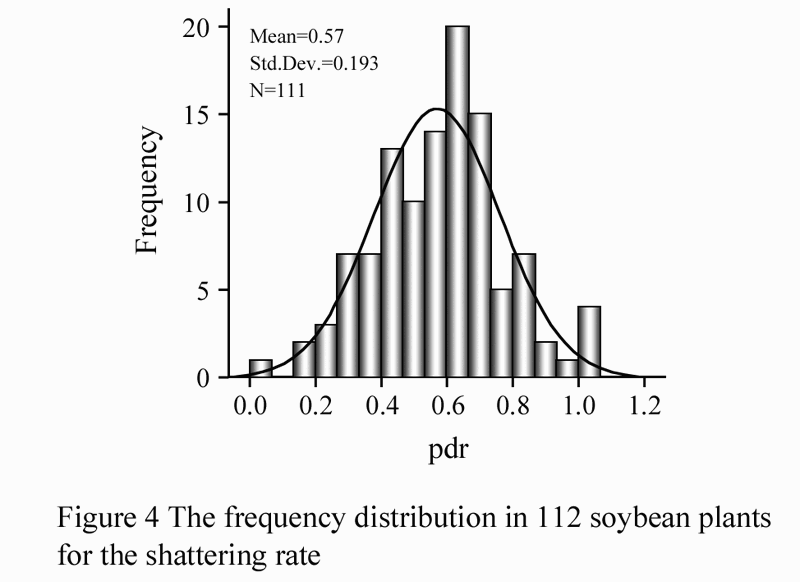 Figure 4 The frequency distribution in 112 soybean plants for the shattering rate |
3.4.2 Phenotype assessment of RTW
Pods were collected (10 pods per plant) when the plant was grown to R7 stage (when one normal pod at any node on the main stem has reached its mature brown or tan pod color). Next, we measured the total width and total thickness of these pods. Measurement of the total width of pods was based on the method of Liu et al., (2003), which measures the width by lining up 10 pods side by side according to the direction of its width, measuring the total width and calculating the average. Measurement of the total thickness of pods was modeled after the above method. A simple device was made by the author, which stacked the pods in accordance with the direction of its thickness. We measured the total thickness, and calculated the average from the total. Then, we calculated the ratio of total thickness to width, and averaged every three repeats. The frequency distribution was obtained with SPSS 16.0 (Figure 5).
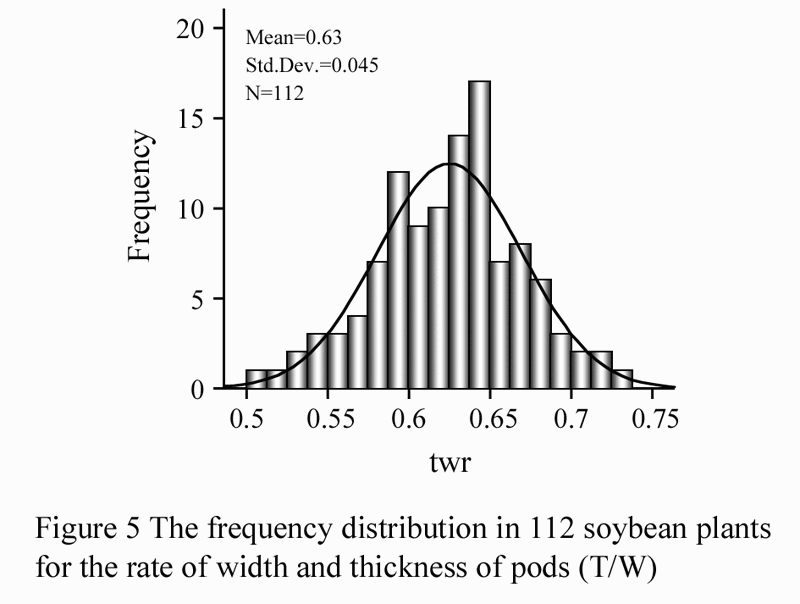 Figure 5 The frequency distribution in 112 soybean plants for the rate of width and thickness of pods (T/W) |
3.4.3 Phenotype assessment of DFM
DFM was the number of days between planting and reaching R8 stage, and averages were obtained from every three repeats. The frequency distribution was obtained with SPSS 16.0 (Figure 6).
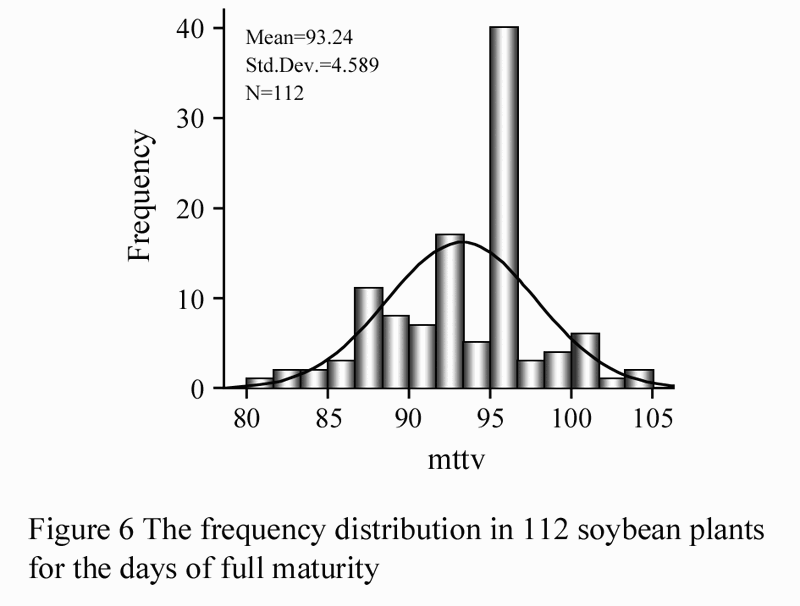 Figure 6 The frequency distribution in 112 soybean plants for the days of full maturity |
Author's contribution
RYL designed and conducted this experiment, and she also completed and modified the first draft. PTG was in charge of experimental design, data analysis, writing and modification of the manuscript. XYL participated in the experimental design and provided instructions on the application of materials. DGZ was the project leader.
Acknowledgement
This research was supported by Scientific Supporting Project of Ministry of Science and Technology of China (2007BAD59B05). Authors (We?) appreciate Dr. XJ Fang from Hainan Institute of Tropical Agricultural Resources (HITAR) for his critical advices, and his reading and reviewing on this paper. We would also like to thank technicians, ZGL, from HITAR for planting and caring for the populations. Lastly, we want to thank the two anonymous reviewers for their useful critical comments and advices on this paper.
References
Agrawal A.P., Basarkar P.W., Salimath P.M., and Patil S.A., 2002, Role of cell wall-degrading enzymes in pod-shattering process of soybean, Glycine max (L.) Merrill, Current Science, 82(1): 58-61
Arnaud N., Girin T., Sorefan K., Fuentes S., Wood T.A., Lawrenson T., Sablowski R., and Østergaard L., 2010, Gibberellins control fruit patterning in Arabidopsis thaliana, Genes and Development, 24: 2127-2132
http://dx.doi.org/10.1101/gad.593410
Arnaud N., Lawrenson T., Østergaard L., and Sablowski R., 2011, The same regulatory point mutation changed seed-dispersal structures in evolution and domestication, Curr. Biol., 21(14): 1215-1219
http://dx.doi.org/10.1016/j.cub.2011.06.008 PMid:21737279
Bailey M.A., Mian M.A.R., Carter Jr. T.E., Ashley D.A., and Boerma H.R., 1997, Pod dehiscence of soybean: Identification of quantitative trait loci, The Journal of Heredity, 88: 152-154
http://dx.doi.org/10.1093/oxfordjournals.jhered.a023075
Caviness C.E., 1969, Heritability of pod dehiscence and its association with some agronomic characters in soybeans, Crop Science, 9: 207-209
http://dx.doi.org/10.2135/cropsci1969.0011183X000900020029x
Christiansen L.C., Dal Degan F., Ulvskov P., and Borkhardt B., 2002, Examination of the dehiscence zone in soybean pods and isolation of a dehiscence-related endopolygalacturonase gene, Plant Cell and Environment, 25: 479-490
http://dx.doi.org/10.1046/j.1365-3040.2002.00839.x
Fry S.C., Smith R.C., Renwick K.F., Martin D.J., Hodge S.K., and Matthews K.J., 1992, Xyloglucan endotransglycosylase, a new wall-loosening enzyme activity from plants, Biochem. J., 282: 821-828
PMid:1554366 PMCid:1130861
Funatsuki H., Ishimoto M., Tsuji H., Kawaguchi K., Hajika M., and Fujino K., 2006, Simple sequence repeat markers linked to a major QTL controlling pod shattering in soybean, Plant Breeding, 125(2): 195-197
http://dx.doi.org/10.1111/j.1439-0523.2006.01199.x
Funatsuki H., Hajika M., Hagihara S., Yamada T., Tanaka Y., Tsuji H., Ishimoto M., and Fujino K., 2008, Confirmation of the location and the effects of a major QTL controlling pod dehiscence, qPDH1, in soybean, Breeding Science, 58: 63-69
http://dx.doi.org/10.1270/jsbbs.58.63
Gong P.T., Mu J.G., Zhao J.R., Wang X.L., Bai Y.N., and Fang X.J., 2006, An integrated soybean genetic linkage map comprising 315 SSRs and 40 AFLPs, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 4(3): 309-316
Kang S.T., Kwak M., Kim H.K., Choung M.G., Han W.Y., Baek I.Y., Kim M.Y., Van K., and Lee S.H., 2009, Population-specific QTLs and their different epistatic interactions for pod dehiscence in soybean [Glycine max (L.) Merr.], Euphytica 166: 15-24
http://dx.doi.org/10.1007/s10681-008-9810-6
Liu B.H., Fujita T., Yan Z.H., Sakamoto S., Xu D.H., and Abe J., 2007, QTL mapping of domestication-related traits in soybean (Glycine max), Annals of Botany, 100: 1027-1038
http://dx.doi.org/10.1093/aob/mcm149 PMid:17684023 PMCid:2759197
Liu X.Y., Wan Y.S., Wang Z., Ma J.K., Shi H., Ren X.J., Ren D.L., Zhang X.H., Li Y.Y., Feng F.M., Xu J., Li Y.Q., and Fang X.J., 2003, Development of soybean recombinant inbred lines Jinf (Jinbean23×ZDD2315) and evaluation by SSR molecular marker and agronomic traits, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 1(2): 157-177
Petersen M., Sander L., Child R., van Onckelen H., Ulvskov P., and Borkhardt B., 1996, Isolation and characterisation of a pod dehiscence zone-specific polygalacturonase from Brassica napus, Plant Mol. Biol., 31(3): 517-527
http://dx.doi.org/10.1007/BF00042225 PMid:8790285
Suzuki M., Fujino K., Nakamoto Y., Ishimoto M., and Funatsuki H., 2010, Fine mapping and development of DNA markers for the qPDH1 locus associated with pod dehiscence in soybean, Molecular Breeding, 25: 407-418
http://dx.doi.org/10.1007/s11032-009-9340-5
Tsuchiya T., 1987, Physiological and genetic analysis of pod shattering in soybean, Japan Agricultural Research Quarterly, 21: 166-175
Tukamuhabwa P., Rubaihayo P., and Dashiell K.E., 2002, Genetic components of pod shattering in soybean, Euphytica, 125: 29-34
http://dx.doi.org/10.1023/A:1015711421223
Wan Y.S., Wang Z., Xiao Y.H., Lv B., and Fang X.J., 2005, A soybean genetic linkage map comprising of 227 SSR loci in a soybean RIL population, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 3(1): 15-20
Yamada T., Funatsuki H., Hagihara S., Fujita S., Tanaka Y., Tsuji H., Ishimoto M., Fujino K., and Hajika M., 2009, A majior QTL, qPDH1, is commonly involved in shattering resistance of soybean cultivars, Breeding Science, 59: 435-440
http://dx.doi.org/10.1270/jsbbs.59.435



. PDF(384KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Ruye Luo
. Pengtao Gong
. Degang Zhao
. Xueyi Liu
Related articles
. Glycine max L.
. Pod dehiscence
. Ratio of thickness to width of pods
. Days of full maturity
. QTL mapping
Tools
. Email to a friend
. Post a comment