Welcome to CropSci Publisher
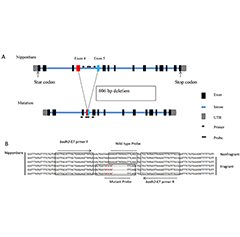
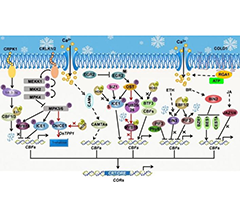
 GWAS Reveals Progress in Genes Related to Rice...
GWAS Reveals Progress in Genes Related to Rice...- Response Analysis of Root and Leaf Physiology ...
- Genetic Regulation of Diurnal Flowering Time D...
- Development of CRISPR-Cas9 Multiple Editing Sy...
- The Genetic Basis of Nutritional Quality Trait...
- Unraveling Key Genetic Factors in Corn Quality...
- Key Genetic Markers Discovered through GWAS in...
- Physiological Mechanisms of Photosynthesis and...
- Nodulation begins with the initiation of infect...(22 Mar.2024)
- Auxin plays a pivotal role in the co-evolution of pl...(22 Mar.2024)
- Auxin plays a pivotal role in the co-evolution of pl...(21 Mar.2024)
- Polyploidization is a major driver of genome diversi...(21 Mar.2024)