Research Report
Microsatellite markers linked to sterility mosaic disease resistance in pigeonpea (Cajanus cajan L. Millsp.) 
2 ICAR-Indian Institute of Pulses Research, Kanpur-208 024, India
3 University of Agricultural Sciences, Dharwad- 580 005, India
 Author
Author  Correspondence author
Correspondence author
Legume Genomics and Genetics, 2016, Vol. 7, No. 6 doi: 10.5376/lgg.2016.07.0006
Received: 09 Mar., 2016 Accepted: 11 Apr., 2016 Published: 12 Jun., 2016
Prakash G. Patil, Byregowda M., Bhuvaneshwara R. Patil, Alok Das, Mary Reena G.A., Sowjanya M.S., and Shashidhar H.E., 2016, Microsatellite markers linked to sterility mosaic disease resistance in pigeonpea (Cajanus cajan L. Millsp.), Legume Genomics and Genetics, 7(6): 1-9 (10.5376/lgg.2016.07.0006)
Sterility mosaic disease (SMD) is a major viral disease of pigeonpea in southern India. In the present study, simple sequence repeat (SSR) markers were employed to identify those genomic regions associated with resistance to Bengaluru isolate of SMD through bulked segregant analysis (BSA) approach. Eighty four F6 recombinant inbred lines (RILs) derived from the cross ICP 8863 (susceptible) x BRG 3 (resistant) were used. A total of 50 hypervariable pigeonpea specific SSR primers (AHSSR) were screened for parental polymorphism study. Out of 50 primers surveyed, 40 primers (80%) were found polymorphic between parents. Based on BSA, three SSR markers viz., AHSSR 50150, AHSSR 34130 and AHSSR 20125 were successfully differentiated resistant and susceptible bulks, and also showed a clearcut differentiation between resistant and susceptible RIL individuals used for constituting these bulks. Segregation analysis of these three SSR markers on 84 F6 RILs exhibited expected Mendelian segregation ratio of 1:1. Single marker analysis (SMA) showed that AHSSR 50150, AHSSR 34130 and AHSSR 20125 were closely associated with SMD resistance and explained 21, 8 and 6 per cent of phenotypic trait variations. The markers identified from this could be useful for SMD resistance breeding in pigeonpea.
Introduction
Pigeonpea (Cajanus cajan (L.) Millsp.) is a major grain legume crop of India, which accounts for 90 per cent of the global pigeonpea production with area 3.86 mha and production of 3.29 mt (FAOSTAT, 2014). Despite of larger area under pigeonpea in India, the production levels are low because of prevalence of various biotic and abiotic stresses. Sterility mosaic disease (SMD) caused by Pigeonpea sterility mosaic virus (PPSMV) (Kumar et al., 2000; Jones et al., 2004) is considered to be major biotic constraint in pigeonpea. It accounts for annual economic losses >US$ 300 million in Indian and Nepal. The virus belongs to genus Emaravirus (Patil and Kumar, 2015) and transmitted by eriophyid mite (Aceria cajani, Channabasavanna) (Kulkarni et al., 2002). The disease is characterized by bushy appearance of plant, mottling of leaves and partial or complete cessation of reproductive structures (Kumar et al., 2003).
The occurrence of five different strains of PPSMV were reported in India (Reddy et al., 1993). Among them, three distinct strains such as Patancheru and Coimbatore were reported as mild, whereas Bengaluru isolate as the most virulent (Kulkarni et al., 2003). Conventional breeding for SMD resistance in pigeonpea has met with limited success. The interplay of host-pathogen-vector interactions with large genotype into environmental interactions has been attributed to limited success. Therefore, deployment of molecular markers in understanding of genetic relationships in available germplasm in relation to traits of interest is an essential prerequisite for any crop improvement programme. Identification of DNA markers tightly linked to genes/quantitative trait loci (QTL) controlling SMD resistance would facilitate enhanced pace and efficiency of breeding pigeonpea for SMD resistance. Literature on DNA markers used for tagging/mapping genes/QTL controlling SMD resistance in pigeonpea is limited. Prasanthi et al. (2009) reported one RAPD marker OPA18800 linked to SMD resistance. Similarly, four AFLP markers linked in coupling phase to the susceptible dominant allele was reported earlier (Ganapathy et al., 2010). Gnanesh et al. (2011a) reported two QTL conferring resistance to SMD using F2 mapping population. There is no report on validation and use of these markers for MAS. Therefore, the current study was undertaken to identify new microsatellite markers which are linked to SMD resistance using RIL population.
1 Results
1.1 Phenotyping of RIL population for SMD resistance
The SMD incidence in RIL population ranged from 0.00 to 100 per cent with a mean of 78.33 per cent. Out of 84 RILs, 4.76 per cent of RILs were resistant, 3.57 per cent were moderately resistant and 91.67 per cent of the RILs were susceptible (Table 1). The resistant RILs showed varying degree of resistance with per cent disease incidence of 0.00 to 8.00 per cent. The RILs No. 1, 17, 62 and 79 showed resistance, RILs No 20, 63 and 81 showed moderately resistant and remaining 77 RILs showed susceptible response. The significance of inbred lines grouped based on SMD reactions were tested by ANOVA. The calculated ‘F’ value was highly significant at 1%, suggesting that the genotypes under consideration differed extensively (Table 2). The frequency distribution graph of RILs for per cent disease incidence revealed negatively skewed (Skewness= - 1.124) and platykurtic (Kurtosis= - 0.019) (Figure 1).
.png) Table 1 Table 1: Classification of RILs into pigeonpea SMD infection response groups. Note: Figures in the parenthesis indicate % SMD incidence of the RILs |
.png) Table 2 Table 2: Analysis of variance of SMD response groups in 84F6 RIL population derived from a cross ICP 8863 x BRG 3. Note: ** Significant at 1 % |
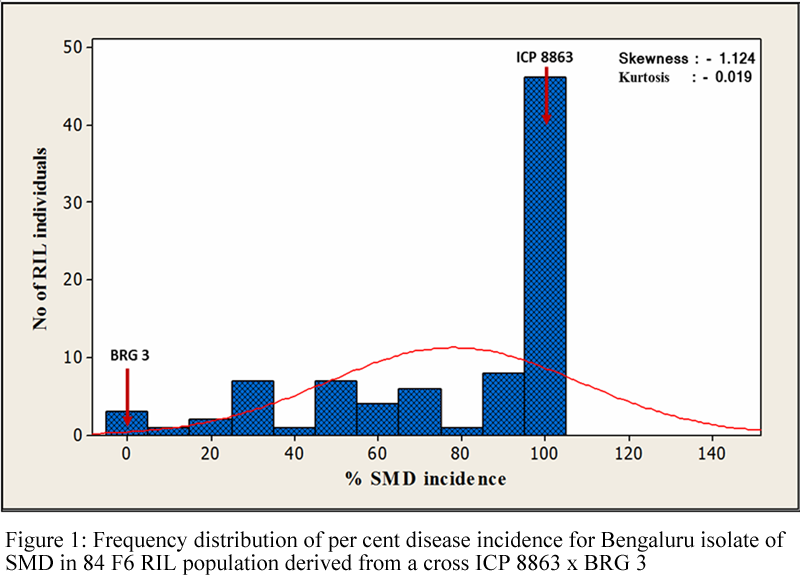 Figure 1 Frequency distribution of per cent disease incidence for Bengaluru isolate of SMD in 84 F6 RIL population derived from a cross ICP 8863 x BRG 3 |
1.2 Parental polymorphism study and bulked segregant analysis
Fifty reported hypervariable pigeonpea specific AHSSR primers (Singh et al., 2012) were surveyed for parental polymorphism between BRG 3 and ICP 8863 (Supplementary Table I). Out of 50 primers screened, 40 (80%) primers were polymorphic between ICP 8863 (S) and BRG 3 (R). This indicated that these primers were highly informative for mapping studies. Out of 40 polymorphic SSR primers, 10 primers could differentiate SMD resistant (consisting of four resistant RILs 1, 17, 62 and 79) and susceptible bulks (four susceptible RILs 7, 9, 11 and 30) (Figure 2a). Out of these, three SSR primers AHSSR 50, AHSSR 34 and AHSSR 20 showed a clearcut differentiation between four resistant and four susceptible RIL individuals used for constituting resistant and susceptible bulks.
1.3 Segregation ratio and single marker analysis
The three SSR markers viz., AHSSR 50150, AHSSR 34130 and AHSSR 20125 obtained from bulked segregant analysis were subjected to analysis of 84 F6 RILs to analyze the segregation patterns of markers (Figure 2b). Chi-square test was performed to examine the goodness of fit of these three SSR markers against Mendelian 1:1 segregation ratio in RILs. All the three SSR marker loci viz., AHSSR 50150, AHSSR 34130 and AHSSR 20125 exhibited Mendelian expected segregation ratio of 1:1 (Table 3). In-order to identify marker trait association, single-marker analysis was performed by taking difference between phenotypic means of each marker classes as an estimate of the phenotypic effects (R2) for different marker genotypes. SMA results showed AHSSR 50150, AHSSR 34130 and AHSSR 20125 were closely associated with SMD resistance and these explained 21, 8 and 6 per cent of phenotypic trait variations (Table 4).
.png) Table 3 Segregation behavior of SSR markers linked to SMD resistance in RIL population of cross ICP 8863 x BRG 3 |
.png) Table 4 Single marker analysis and identification of markers associated with SMD resistance in RILs of cross ICP 8863 x BRG 3. Note: * Significant at 5 %, ** Significant at 1 % |
.png) Figure 2 BSA analysis and segregation pattern of SSR marker AHSSR 50150 linked to SMD resistance in RIL population of cross ICP 8863 x BRG 3. Note: Where L=100bp ladder, 2a- BSA analysis, 1-BRG 3 (Resistant), 2-ICP 8863 (Susceptible), 3- resistant bulk, 4- susceptible bulk, 2b- segregation of linked marker, P1-BRG 3 (R), P2-ICP 8863(S),1-84 RIL individuals |
2 Discussion
2.1 Reaction of RIL population to SMD
The 84 F6 RILs were grouped into three classes based on per cent disease incidence viz., resistant with no apparent symptoms, moderately resistant with partial symptoms and susceptible with severe mosaic symptoms based on the standard scale (Singh et al., 2003). SMD incidence in 84 RILs ranged from 0.00 to 100 per cent with mean of 78.33 per cent. One-way analysis of variance (ANOVA) indicated significant differences between the three SMD infection response groups. Similarly, the means for SMD response groups of TTB 7 × ICP 7035 derived F2:3 progenies were found significant at 1 per cent against Bengaluru isolate (Gnanesh et al., 2011a). In RIL population large number of plants could be classified into categories of susceptible class followed by few moderately resistant and resistant groups. These results indicated that for Bengaluru isolate susceptibility is controlled by dominant genes, therefore number of plants with high level of resistance to SMD is fewer in F6 RIL population. Resistance to SMD in general is controlled by recessive genes and the causal organism has higher level of virulence, hence it is very difficult to realize plants with resistance to SMD in Bengaluru. The frequency distribution graph for SMD revealed platykurtic (Kurtosis= -0.019) and negatively skewed (Skewness= -1.124) indicating resistance is due to duplicate gene interaction involving two or more genes. Positive skewness is caused by complementary gene interactions, while negative skewness is caused by duplicate gene interactions predominantly in the same directions (Snape and Riggs, 1975). Gnanesh et al. (2011b) studied inheritance of resistance to Bengaluru isolates in F3 families of cross ICP 7035 (R) x TTB 7 (S) and found absence of resistant plants indicating action of two or more genes in controlling resistance. Similarly, resistance to SMD has been also been reported to be oligogenic in nature (Singh et al., 1983; Sharma et al., 1984; Balu and Rathnasamy, 2003; Nagaraj et al., 2004; Ganapathy, 2009).
2.2 Bulked segregant analysis and marker trait association
The combination of bulked segregant analysis (Michelmore et al., 1991) and highly polymorphic PCR based markers permits the identification of useful molecular markers for breeding programme. In this study, we found out of 50 AHSSR primers screened for parental polymorphism, 40 (80%) primers were polymorphic. This confirmed these are hyper variable pigeonpea specific SSR markers with high polymorphism as reported (Singh et al., 2012). Polymorphic AHSSR primers and parents used for development of RIL population were subjected to bulked segregant analysis. Out of 40 polymorphic SSR markers, three SSR marker loci viz., AHSSR 50150, AHSSR 34130 and AHSSR 20125 were found bulk positive and these differentiated most of the individuals used for constituting resistant and susceptible bulks. The genotypic data of each primers for the RILs along with SMD incidence data were used for markers trait association through single marker analysis. Similarly, four AFLP markers each for resistance to SMD and fusarium wilt were identified in pigeonpea through bulked segregant analysis (Ganapathy, 2009; Ajay, 2010).
In single-marker analysis, each marker is considered as treatment with two marker classes for RIL mapping population and phenotypic traits considered as dependent variables. This analysis involved comparing for each SSR marker, the phenotypic means of the two marker classes. The difference between the phenotypic means of each marker classes was used as an estimate of the phenotypic effects (R2) of different marker genotypes. Single-marker analysis results showed AHSSR 50150, AHSSR 34130 and AHSSR 20125 were closely associated with SMD resistance and these explained 21, 8 and 6 per cent of phenotypic variation respectively. Similarly, single marker analysis approach was used to associate markers with disease resistance in other crops viz., cassava mosaic disease resistance in cassava (Lokko et al., 2005) and yellow mosaic virus resistance in cowpea (Gioi et al., 2012).
3 Conclusion
The SSR markers identified in this study needs to be validated in other genetic backgrounds of pigeonpea. These markers will be useful for indirect selection of resistant pigeonpea genotypes in early segregating generations and marker assisted backcross breeding for SMD resistance.
4 Materials and Methods
4.1 Plant material
The material for the present study comprised of eighty four F6 recombinant inbred lines (RILs) derived from the cross of two diverse genotypes such as SMD resistant parent (BRG 3) and susceptible parent (ICP 8863). The seeds of these lines were obtained from All India Coordinated Research Project on Pigeonpea, UAS, Bengaluru, India.
4.2 Phenotyping for SMD resistance
The seeds of 84 F6 RILs having minimum of ten seedlings per family along with their parents were raised in the polythene bags during kharif-2014 at experimental plots of ZARS, UAS, Bengaluru (Supplementary Table II). Phenotyping against Bengaluru isolate of SMD was carried out following “Leaf stapling method” as described (Nene et al., 1981). Fifteen days old seedlings were selected to staple SMD infected leaves. Disease incidence was scored visually by counting healthy and diseased plants for each genotype at 15 days intervals up to 60 days. Per cent disease incidence (PDI) was calculated by using formula as described earlier (Singh et al., 2003). The recombinant inbred lines were grouped as resistant (0–10% of plants infected); moderately resistant (10.1–30% of plants infected) and susceptible (30.1–100% of plants infected).
.png)
The analysis of variance (ANOVA) for SMD scoring on RILs was performed to test the significance of differences between inbred lines. The per cent disease incidence values of RILs were also used to estimate coefficients of skewness and kurtosis using SPSS 16.0 software (SPSS Inc., Chicago, USA).
4.3 Collection of leaf samples and DNA extraction
All the 84 RILs including parents were labeled and leaf samples were collected before stapling the infected leaves. Genomic DNA was extracted from the young leaves of 15 days old seedlings using a modified cetyl trimethyl ammonium bromide method (Agbagwa et al., 2012). Finally, DNA were quantified by both visual and UV spectrophotometer and diluted to a concentration of 10 ng/μl.
4.4 SSR analysis
Fifty reported hyper variable pigeonpea specific AHSSR primers (Singh et al., 2012) were screened to identify polymorphism between the resistant (BRG 3) and susceptible (ICP 8863) parents. Polymerase chain reaction was performed in 10 μl reaction mixture constituting 10 ng genomic DNA, 1μl 10X PCR buffer (15 mM of MgCl2), 1 mM dNTP mix, 10 μM of forward and reverse primer and 0.2 μl Taq DNA polymerase (3U/ μl). The PCR reactions were conducted in Thermocycler (Eppendorf, USA) for each primer set using the following reaction conditions: initial denaturation at 94ºC for 5 min, followed by 35 cycles of 94ºC for 1 min, 55ºC for 1 min, 72ºC for 1 min and final extension of 72ºC for 7 min. All the PCR amplicons were separated on 6% horizontal PAGE (Patil et al., 2015). Gel was visualized and photographed in gel documentation unit (Alpha Innotech, UK). The bands generated by SSR primers were scored by giving code ‘1’ for those RIL individuals showing size similarity with ICP 8863, ‘3’ for those showing band size similarity with BRG 3 and ‘2’ for those showing similar banding pattern with both parents.
4.5 Bulked segregant analysis and association of markers
Bulked segregant analysis was carried out essentially as described by Michelmore et al. (1991) using polymorphic SSR markers. The resistant and susceptible DNA bulks were prepared from RIL individuals by pooling aliquots containing equal amount of DNA (0.25 μg) from each of the four resistant and susceptible individuals based on response to SMD. The putatively linked SSR markers identified from BSA were used to screen on individuals used to constitute respective bulks. The SSR markers which could differentiate individuals of each bulk were finally screened on entire 84 RIL individuals. The associations of SSR markers with SMD resistance trait were analyzed with single marker analysis using SPSS 16.0 software (SPSS Inc., Chicago, USA). Chi-square test (χ2) was performed to examine the goodness of fit between the expected Mendelian ratios for the segregation data of linked SSR markers.
Authors' contributions
PGP - Conception and design, acquisition of data, analysis and drafting the manuscript
MB - Substantial contributions to conception, design and providing plant materials
BRP- Sharing of primers and helped in drafting the manuscript and revising it critically
AD - Helped in drafting the manuscript and revising it critically
MGA - Participated acquisition of data and statistical analysis
SMS - Statistical analysis and drafting manuscript
HES - Supervised, guided, helped in interpretation of data and given final approval for communication
Acknowledgement
Authors would like acknowledge the University of Agricultural Sciences (UAS), GKVK, Bengaluru, India for providing all the logistical support to carry out this research work.
Agbagwa I.O., Datta S., Patil P.G., Singh P., and Nadarajan N., 2012, A protocol for high-quality genomic DNA extraction from legumes, Genet. Mol. Res, 11: 4632-4639
http://dx.doi.org/10.4238/2012.September.14.1
Ajay B.C., 2010, Identification of AFLP markers linked to fusarium wilt resistance and genetic variability studies in pigeonpea (Cajanus cajan (L.) Millsp.), Ph.D Thesis, University of Agricultural Sciences, Bengaluru
Balu A. P., and Rathnasamy R., 2003, Inheritance to sterility mosaic disease in pigeonpea (Cajanus cajan (L.) Millsp), Crop Res, 25 (2): 301-304
FAOSTAT, 2014, Food and Agriculture Organization of United Nations, http://faostat.fao.org
Ganapathy K.N., 2009, Genetic analysis and identification of DNA markers linked to sterility mosaic disease resistance in pigeonpea [Cajanus cajan (L.) Millsp.], PhD Thesis, University of Agricultural Sciences, Bengaluru
Ganapathy K.N., Byregowda M., Venkatesh S.C., Ramachandra R., Gnanesh B.N., and Girish G., 2010, Identification of AFLP markers linked to sterility mosaic disease in pigeonpea [Cajanus cajan (L.) Millsp.], Inter. J. Integ. Biotechnol, 7 (13): 145-149
Gioi T.D., Boora K.S., and Chaudhary K., 2012, Identification and characterization of SSR markers linked to yellow mosaic virus resistance gene(s) in cowpea (Vigna unguiculata), Inter. J. Plant Res, 2(1): 1-8
http://dx.doi.org/10.5923/j.plant.20120201.01
Gnanesh B.N., Bohra A., Sharmaa M., Byregowda M., Pandea S., Wesleya V., Saxena R.K., Saxena K.B., Kavikishor P.B., and Varshney R.K., 2011a, Genetic mapping and quantitative trait locus analysis of resistance to sterility mosaic disease in pigeonpea (Cajanus cajan (L.) Millsp.), Field Crops Res, 54 (70): 1-9
http://dx.doi.org/10.1016/j.fcr.2011.04.011
Gnanesh B.N., Ganapathy K.N., Ajay B.C., and Byre Gowda M., 2011b, Inheritance of sterility mosaic disease resistance to Bangalore and Patancheru isolates in pigeonpea (Cajanus cajan (L.) Millsp.), Electronic J. Plant Breed, 2 (2): 218-223
Jones A.T., Kumar L.P., Saxena K.B., Kulkarni N.K., Muniyappa V., and Waliyar F., 2004, Sterility mosaic disease the “Green Plague” of pigeonpea: advances in understanding the etiology, transmission and control of a major virus disease, Plant Dis, 88: 436-445
http://dx.doi.org/10.1094/PDIS.2004.88.5.436
Kulkarni N.K., Kumar P.L., Muniyappa V., Jones A. T., and Reddy D.V.R., 2002, Transmission of pigeonpea sterility mosaic virus by the eriophyid mite, Aceria cajani (Acari: Arthropoda), Plant Dis, 86: 1297-1302
http://dx.doi.org/10.1094/PDIS.2002.86.12.1297
Kulkarni N.K., Reddy A.S., Kumar L.P., Vijaynarasimha J., Rangaswamy K.T., Muniyappa V., Reddy L.J., Saxena K.B., Jones A.T., and Reddy D.V.R., 2003, Broad-based resistance to pigeonpea sterility mosaic disease in accessions of Cajanas scarabaeoides (L.) Benth, Indian J. Plant Prot, 31: 6-11
Kumar P.L., Jones A.T., and Reddy D.V.R., 2003, A novel mite transmitted virus with a divided RNA genome closely associated with pigeonpea sterility mosaic disease. Phytopath., 93: 71-81
http://dx.doi.org/10.1094/PHYTO.2003.93.1.71
Kumar P.L., Jones A.T., Sreenivasulu P., and Reddy D.V.R., 2000, Breakthrough in the identification of the causal agent of pigeonpea sterility mosaic disease, J. Mycol. Plant Pathol, 30 (2): p.249
Lokko Y., Danquah E.Y., Offei S.K., Dixon A.G.O., and Gedil M.A., 2005, Molecular markers associated with a new source of resistance to the cassava mosaic disease, African J. Biotechnol, 4 (9): 873-881
Michelmore R.W., Paran I., and Kesseli R.V., 1991, Identification of markers linked to disease resistance genes by bulked segregant analysis: A rapid method to detect markers in specific genomic regions by using segregating populations, Proc. Natl. Acad. Sci, 88: 9828-9832
http://dx.doi.org/10.1073/pnas.88.21.9828
Nagaraj K.M., Chikkadevaiah, and Kulkarni R.S., 2004, Inheritance of resistance to sterility mosaic virus in pigeonpea (Cajanus cajan (L.) Millsp.), Indian J. Genet, 64 (2): 118-120
Nene Y.L., Kannaiyan J., and Reddy M.V., 1981, Resistance to major pigeonpea diseases, In: Proc. Int. Workshop on pigeonpea, 15-19, Dec. 1980, ICRISAT, Patancheru, India, 1121-1128
Patil B.L., and Kumar P.L., 2015, Pigeonpea sterility mosaic virus: a legume-infecting Emaravirus from South Asia, Mol. Plant Pathol, 13: doi:10.1111/mpp.12238.
http://dx.doi.org/10.1111/mpp.12238
Patil P.G., Byre Gowda M., Kundur P.J., Vimarsha H.S., and Shashidhar H.E., 2015, Upgraded horizontal polyacrylamide gel units for DNA marker genotyping, Indian Journal of Science and Technology, 8(9): 822-827
http://dx.doi.org/10.17485/ijst/2015/v8i9/56638
Prasanthi L.R., Reddy B.V.B., Rani K.R., Sivaprasad Y., Rajeswari T., and Reddy K.R., 2009, RAPD and SCAR marker linked to the sterility mosaic disease resistance gene in pigeonpea (Cajanus cajan L. Millsp.), The Asian and Australian Journal of Plant Science and Biotechnology, 3(1): 16-20
Reddy M.V., Raju T.N., Nene Y.L., Ghanekar A.M., Amin K.S., Arjunan G., Astaputre J.Y., Sinha B.K., Muniyappa Y., Reddy S.Y., Gupta R.P., and Gangadharan K., 1993, Variability in sterility mosaic pathogen of pigeonpea in India, Indian Phytopath, 37-43
Sharma D., Gupta S.C., Rai G.S., and Reddy M.V., 1984, Inheritance of resistance to sterility mosaic disease in pigeonpea, Indian J. Genet, 44 (1): 84-90
Singh A.P., Singh U.P., Singh R.M., and Raina R., 2003, Relative efficiencies of four population improvement schemes in pigeonpea (Cajanus cajan (L.) Millsp.), Crop Res, 25 (2): 305-311
Singh B.V., Pandya B.P., Gautam P.L., Beniwal S.P.S., and Pandey M.P., 1983, Inheritance of resistance to sterility mosaic virus in pigeonpea, Indian J. Genet, 43: 487-493
Singh N.K., Gupta D.K., Jayaswal P.K., Mahato A.K., Dutta S., Singh S., Bhutani S., Dogra V., Singh B.P., Kumawat G., Pal J.K., Pandit A., Singh A., Rawal H., Kumar A., Prashat G. R., Khare A., Yadav R., Raje R.S., Singh M.N., Datta S., Fakrudin B., Wanjari K.B., Kansal R., Dash P.K., Jain P.K., Bhattacharya R., Gaikwad K., Mohapatra T., Srinivasan R., and Sharma T.R., 2012, The first draft of the pigeonpea genome sequence, J. Plant Biochem. Biotechnol, 21: 98-112
http://dx.doi.org/10.1007/s13562-011-0088-8
Snape J.W., and Riggs T.J., 1975, Genetical consequences of single seed descent in the breeding of self pollinating crops, Heredity, 35: 211-219



. PDF(176KB)
. FPDF
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Prakash G. Patil
. Byregowda M.
. Bhuvaneshwara R. Patil
. Alok Das
. Mary Reena G.A.
. Sowjanya M.S.
. Shashidhar H.E.
Related articles
. Pigeonpea
. Single marker analysis
. Sterility mosaic disease
. SSR markers
Tools
. Email to a friend
. Post a comment